Struktura czaszek zwierząt. Struktura mózgu zwierząt i opon mózgowych. Tętnice mózgowe
GŁOWICA SZELETONOWA. CZASZKA
Pojawienie się głowy i jej szkieletu wiązało się z komplikacją ogólnej organizacji przodków kręgowców, występującą w walce o byt i ze względu na postępujący rozwój ciała, dostosowującego się do zmieniających się warunków życia. W przedniej części dwubiegunowego ciała zwierzęcia rozwijają się dwie niezależne części szkieletu głowy (ryc. 32). Jeden jest urządzeniem ochronnym dla rozwijającego się mózgu - czaszki - czaszki, drugi (sekcja twarzy) - w związku z ulepszaniem urządzeń do lepszego, lepszego wychwytywania żywności i jej pierwotnego przetwarzania. Tak więc na przednim końcu dolnej części trzonu kręgowego powstały dwie części szkieletu głowy: czaszka, która tworzy jamę czaszki dla mózgu, i sekcja trzewna, która dalej tworzy twarz - zanika. Te dwa oddziały połączyły ogólną lokalizację i fakt, że ruchomy aparat szczękowy sekcji twarzy znalazł podparcie dla ich mięśni na czaszce.
Mózg delfinów jest drugim najpotężniejszym i najbardziej złożonym mózgiem zwierząt. Inteligencję można zdefiniować jako miarę zdolności mózgu do przetwarzania informacji w sposób, który rozwiązuje problemy i zwiększa przeżycie. Pomiar inteligencji może być zupełnie inny dla zwierząt, które ewoluowały w wodzie, niż zwierząt żyjących na lądzie, ponieważ problemy niezbędne do przetrwania w wodzie są bardzo różne od tych wymaganych na lądzie. Toczy się wiele debat na temat „inteligencji delfinów”.
Duże zwierzęta mózgowe, takie jak ludzie, szympansy i delfiny, mają wiele wspólnego, zwykle żyją długo, żyją w płynnych grupach społecznych, aw dzieciństwie wykazują całkowitą zależność rodzicielską. Dlatego niemożliwe jest pełne oszacowanie poziomu inteligencji tego zwierzęcia. Ale innym sposobem zaproponowanym do pomiaru inteligencji jest określenie miary względnej wielkości mózgu, zdefiniowanej jako stosunek rzeczywistej masy mózgu do przewidywanej masy mózgu zwierzęcia o danym rozmiarze.
Ryc. 32. Czaszka rekina
Każdy dział rozwijał się z różnych elementów znajdujących się w tym obszarze: chrząstki uformowanej po bokach cięciwy i przed nią (czaszka) oraz z pierwszych łuków trzewnych (twarz). Kości pochodzące z tych miejsc przechodzą trzy etapy kostnienia w ontogenezie i są wtórne. Następnie poszczególne części zewnętrznego szkieletu, które tworzą kości szkieletowe czaszki, uczestniczą w tworzeniu czaszki. Kości szkieletowe czaszki są pierwotne, aw ontogenezie tylko dwa etapy kostnienia przechodzą - błoniaste i kostne (ryc. 33).
Ten pomiar zakłada, że \u200b\u200bim wyższa liczba, tym wyższa inteligencja. Ponadto stopień koagulacji kory mózgowej wydaje się miarą inteligencji. Im większa złożona kora, tym więcej miejsca w mózgu, aby pomieścić dodatkowe neurony, za pomocą których można przeprowadzić przetwarzanie informacji. Niedawno opublikowane informacje na temat wzrostu fałdowania kory mózgowej Alberta Einsteina w porównaniu z innymi ludźmi potwierdzają tę teorię. Jedynym zwierzęciem o bardziej złożonej korze niż ludzie jest delfin.
Rozwój śródmózgowia
To zdjęcie dotyczy ludzkiego mózgu. głównym powodem tak dużego mózgu jest przetwarzanie informacji ze złożonej echolokacji delfina lub układu biosonarnego. Ale nie ma dowodów na poparcie tego przekonania. Najczęstszą teorią jest to, że ten większy mózg ewoluował, aby wspierać bardziej złożone zdolności poznawcze. Mogą pamiętać wydarzenia i badać koncepcje, zmieniając swoje zachowanie w wyniku wcześniejszych doświadczeń. Mogą komunikować się ze sobą podczas zachowań kooperacyjnych, zarządzać relacjami w swoich strąkach i wychowywać dzieci.
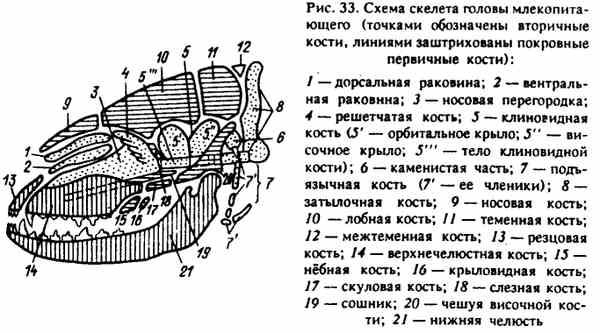
Ryc. 33. Schemat szkieletu głowy ssaka (kropki wskazują kości wtórne, linie pokrywają pierwotne kości powłokowe)
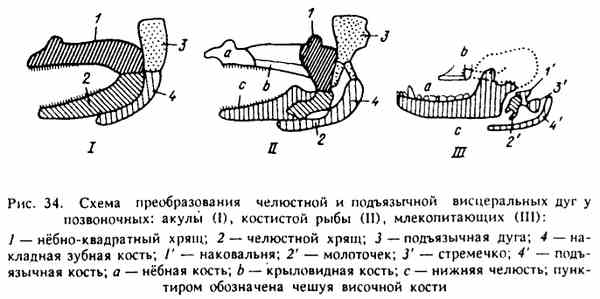
Ryc. 34. Schemat transformacji szczęki i podjęzykowych łuków trzewnych u kręgowców
Rozumieją nie tylko słowa języka symbolicznego, ale także interpretują porządek składniowy języka. To zrozumienie składni bardzo wskazuje na inteligencję. Ruchome gwizdy tworzone przez delfiny dowodzą, że delfiny mają samoświadomość lub zdolność posiadania koncepcji jaźni i wiedzieć, że istnieje ona jako indywidualna istota. Samoświadomość istnieje w przedniej korze czołowej. Oprócz delfinów samoświadomość najwyraźniej istnieje tylko u dużych naczelnych mózgu i ludzi.
W wyniku badania choroby Alzheimera u ludzi stwierdzono, że ludzkie neurony wrzecionowe są związane ze zdolnością rozpoznawania, zapamiętywania, rozumowania, komunikowania się, postrzegania, dostosowywania się do zmian, rozwiązywania problemów i rozumienia. Wybuchy zmarłych pacjentów z chorobą Alzheimera wykazują wysoką częstość występowania zużytych neuronów wrzecionowych. Ostatnio neurony wrzeciona zostały wyizolowane w mózgach prawdziwych wielorybów. Ten wniosek pokazuje, że prawdziwe wieloryby posiadają te zaawansowane umiejętności, które w przeszłości były kojarzone tylko z ludźmi i naczelnymi.
Czaszka została utworzona z wielu kości, które tworzyły oddzielne części czaszki i twarzy. Pomiędzy kościami znajdują się szwy wypełnione gęstym tkanka łączna. Pomiędzy niektórymi kościami w ontogenezie powstają znaczące przestrzenie tkanki łącznej, zwane fontanelami; stają się skostniałe przez starość. Tak więc u ssaków takie fontanele znajdują się między kościami potylicznymi i ciemieniowymi czaszki lub między kością potyliczną, ciemieniową i skroniową.
Ogólna charakterystyka rozwoju mózgu
Od dawna podejrzewa się, że delfiny potrafią rozpoznawać ludzi i przedmioty, zapamiętywać zadania, rozwiązywać problemy, dostosowywać się do zmian i badać złożone zadania. Dolphin Institute w Honolulu na Hawajach bada zachowanie ssaków morskich od 30 lat. Dzięki swojej pracy znacznie poszerzyli naszą wiedzę o zachowaniu delfinów i inteligencji. Poniżej znajduje się krótki przegląd wyników wielu projektów badawczych.
Ogólna charakterystyka struktury mózgu
Delfiny, podobnie jak ludzie, są zdolne do behawioralnego wyrazu twarzy. Innymi słowy, mogą naśladować zachowanie swoich trenerów. Jeśli ktoś podniesie nogę, delfin może rozpoznać związek, jaki ma część ludzkiego ciała z własną anatomią i podniesie ogon. Wskazuje to, że zwierzę może kojarzyć część swojej anatomii z ludzką postacią.
W procesie filogenezy niektóre kości, które budują szkielet głowy niższych kręgowców, zmieniają swoje funkcje. Tak więc prymitywne szczęki ryb zastępowane są przez kości powłokowe, ale kości szczęki wcale nie znikają, zmieniają swoją funkcję i pojawiają się w górnych kręgowcach przez słuchowe kosteczki ucha środkowego (ryc. 34).
U zwierząt lądowych, w związku ze zmianą oddychania skrzelowego na oddychanie płucne, jama nosowa rozwija się na podstawie rowków węchowych na twarzy, komunikując się z przodu wąskim ujściem między siekaczem, kościami szczękowymi i otwieraczem. Dziury te nazwano pierwotnymi hoansami. U zwierząt z pierwotnymi choanami pokarm nie może przejść przez jamę ustną, gdy powietrze przechodzi przez jamę nosową. Pod tym względem struktura jamy nosowej jest ulepszana i skomplikowana. Rozmieszczenie choinek stopniowo się zmienia, a już u gadów otwierają się za jamą nosową do gardła i są nazywane drugorzędowymi choanami.
Może także naśladować innego delfina. Dowodem na to jest zachowanie synchroniczne. Jeśli jeden delfin ma zamiar zrobić ukłon, inne zwierzę może skopiować to zachowanie i jednocześnie skoczyć z tym zwierzęciem. Mówi się, że jeden delfin działa jako „demonstrator”, podczas gdy drugie zwierzę jest „naśladowcą” takiego zachowania.
Delfiny mogą również interpretować zachowania telewizyjne i reagować na gesty wyświetlane na ekranie po raz pierwszy, gdy są wystawione na telewizję. Jest to pierwsza demonstracja jakiejkolwiek reakcji behawioralnej zwierząt na gesty telewizyjne. Psy, koty i szympansy nie wykazały takich odpowiedzi w podobnych protokołach badawczych. Badanie to zmieniło sposób, w jaki klasyfikowaliśmy delfiny przede wszystkim jako specjalistów akustycznych. Teraz rozumiemy, że są oni również specjalistami od wizualizacji, wykorzystując zarówno wzrok, jak i dźwięk, aby wyróżniać się w środowisku wodnym.
Czaszka jest połączona z pierwszym kręgiem szyjnym (atlas) i tworzy jamę czaszki, w której znajduje się mózg. Ta jama czaszki - cavum cranii ma dach, tył, przód, ściany boczne i dno jamy czaszki.
DACH CZASZKI - kalwaria.
U świń, koni i psów tworzy go sparowana kość czołowa - os frontale (z przodu), sparowana kość ciemieniowa - os parietale i niesparowana kość między ciemnymi - os interparietale - (z tyłu). Kość ciemieniowa przylega ogonowo do kości potylicznej. U młodych zwierząt w miejscu szwów między kościami ciemieniowymi i potylnymi powstaje ciemiączka potyliczne, w którym kładzie się sparowane kostnienie kostnienia, z którego powstaje niesparowana kość międzyścienna. Znajduje się w linii środkowej w rogu między dwiema kościami ciemieniowymi i potylicznymi. Można go zobaczyć z wnętrza jamy czaszki w postaci znacznego pogrubienia kości, z którego przegrzebki rozciągają się na prawo i na lewo. Jest to test kostny - tentorium osseum, do którego przyczepiony jest test błoniasty opony twardej.
Wykazano, że delfiny rozpoznają się w lustrze, wykorzystując tak zwane „testy awaryjne” lub wykonując ruchy, badając siebie pod kątem tego ruchu. Odkrycie to jest nieoczekiwane, ponieważ delfiny przede wszystkim doświadczają świata poprzez dźwięk, a ich system echolokacji nie będzie działał w dwuwymiarowym odbiciu.
Koniec mózgu - telencefalon
Są świadomi swoich ostatnich zachowań i mogą powtórzyć zachowanie lub, na żądanie trenera, przeprowadzić zachowanie, które nie zostało ostatnio wykonane. Zespoły reprezentujące „powtórzenie” lub zespoły reprezentujące „dowolne” skutkują odpowiednio powtórzeniem ostatnich zachowań lub wyborem niektórych nierzeczywistych zachowań. To pokazuje zdolność delfina do zachowania zachowania mentalnego, które ostatnio wykonał, i do aktualizacji tego obrazu, gdy każde nowe zachowanie jest wykonywane, powtarzając ostatnie zachowanie w tej sekwencji na żądanie.
U koni, świń i psów przednią część dachu jamy czaszki tworzy przednia kość - os frontale. Jest to łaźnia parowa, reprezentowana przez trzy płyty:
1) górna część czołowa, do której przymocowana jest kość nosowa z przodu;
2) orbital, zstępujący bocznie i uczestniczący wraz ze skrzydłami kości kośćcowej i łuskami kość skroniowa w formacji na zewnątrz czaszki wewnętrznej ściany orbity. (Na styku tych dwóch płytek proces zygomatyczny - zygomatics procesus, który u koni i przeżuwaczy osiąga proces jarzmowy kości skroniowej lub czasowy proces kości jarzmowej i tworzy pełny pierścień orbity u świń i psów, odchodzi poza kość czołową na styku tych dwóch płytek) nie dociera i tworzy niekompletny pierścień. Na podstawie tego procesu można znaleźć otwór oczodołowy - otwór nadoczodołowy.
Delfiny odpowiadają trenerowi wskazującemu obiekt. Delfiny nie tylko rozumieją i odpowiednio reagują na osobę wskazującą bezpośrednio na obiekt, ale reagują odpowiednio na punkt przecięcia. Przykładem odpowiedniej odpowiedzi na wskazanie byłoby pobranie obiektu, na który wskazuje trener, lub przeniesienie obiektu z punktu A do punktu B.
Podobne badanie zdolności delfinów do koncentrowania się na zadaniach wymagających długotrwałej koncentracji przyniosło ciekawe wyniki. To pokazuje zdolność delfina do zachowania czujności przez długi czas i szybkiego rozróżniania krytycznych i niekrytycznych obrazów i dźwięków z dużą dokładnością.
U przeżuwaczy otwór nadoczodołowy znajduje się u podstawy procesu jarzmowego, z którego korytko płynie tam iz powrotem, przechodząc na orbitę do kanału.
U świń dziura przechodzi do rynny, docierając do kości nosowych.
Psy nie mają otworu podoczodołowego.);
3) wewnętrzna pionowa (niewidoczna z zewnątrz), tworząca przód dachu jamy czaszki.
Oprócz samoświadomości delfiny doświadczają podstawowych emocji, uczestniczą w pewnym stopniu abstrakcyjnej, konceptualnej myśli, wybierają swoje działania, uczą się poprzez obserwację, rozumieją strukturę swojego otoczenia, uczą się, co działa, a co nie, rozwiązując problemy. tworzyć nowe rozwiązania problemów, z którymi są prezentowane. Podczas interakcji z osobą wydaje się, że zdaje sobie sprawę z różnicy między dziećmi a dorosłymi, i jest bardziej łagodna i cierpliwa wobec dzieci.
Neuronauka sugeruje, że ludzki mózg jest standardem, według którego należy porównywać mózgi wszystkich zwierząt. Ale wiele zwierząt wykazuje wyjątkowe talenty i wysoce zaawansowane zdolności umysłowe oraz zachowania społeczne. Ich inteligencja jest bardzo różna od człowieka i w pewien sposób go przewyższa. Wiele obszarów badań pokazuje unikalne mózgi zwierząt z wyjątkowymi talentami do odnawiania zwierząt.
Pomiędzy tymi trzema płytkami (czołową, oczodołową i wewnętrzną) w prawej i lewej kości czołowej tworzą się przestrzenie - wnęki wypełnione powietrzem i pokryte błonami śluzowymi. Nazywa się je zatokami lub zatokami. W kości czołowej jest to sparowana zatoka czołowa - zatok czołowy. Zatoki symetryczne są oddzielone przegrodą strzałkową i nie są przekazywane. Wewnątrz zatok nadal znajdują się nieciągłe przegrody, które dzielą każdą zatokę na komórki, które komunikują się ze sobą. Zatoki czołowe przechodzą do ciemieniowego i potylicznego (u świń i bydła).
Słynny artykuł filozoficzny z pytaniem, czy możemy zrozumieć, jak to jest być nietoperzem. W rzeczywistości ludzie nie są w stanie doświadczyć istot o całkowicie odmiennych zmysłach i systemy napędowe. Struktury w mózgach zwierząt są zupełnie inne niż ludzie, a ocena ich funkcji w porównaniu z ludzkim mózgiem nie wyjaśnia bardzo zaawansowanego zachowania.
Większość zwierząt ma znacznie więcej inteligencji, niż można by się spodziewać po prostym anatomicznym porównaniu z ludzkim mózgiem. Obecnie inteligencja słoni, delfinów, wielorybów, ośmiornic i naczelnych jest dobrze znana. Niedawno odkryto, że delfiny przypominają wokalizacje swoich czołgistów sprzed dwudziestu lat - niesamowity wyczyn pamięci. Zaawansowana inteligencja terapeutycznych psów była odkryciem, że wiele zwierząt ma kulturę.
U przeżuwaczy istotną cechą czaszki jest to, że tylko kość czołowa tworzy dach jamy czaszki.
U bydła ma znaczne wymiary, wewnątrz dość silnych zatok, przechodzących w zatoki ciemieniowe i nosowe, a także nadal puste, napalone procesy - processus cornuales, których wnęka komunikuje się z zatokami czołowymi, odchodzą od jego zewnętrznej powierzchni. Dlatego musimy pamiętać, że podczas cięcia rogu wejście do zatoki czołowej otwiera się przez otwór uformowany w procesie pustego rogu kości czołowej. Tylna część sklepienia czaszki tworzy pogrubioną krawędź (elewacja między rogami), z którą łączą się kości ciemieniowe i ciemieniowe od dołu. Jest podobny do grzebienia potylicznego u innych zwierząt. Kość czołowa, tworząca cały dach jamy czaszkowej u bydła, jakby popycha kości ciemieniowe i między ciemne, które u tych zwierząt znajdują się za, nieco wyżej niż kość potyliczna, pod grzebieniem czołowym, do tylnej i bocznej ściany jamy. Kości ciemieniowe wchodzą w ścianę boczną między kościami czołowymi (powyżej) i skroniowymi (poniżej), uzyskując kształt litery P, na której kości czołowej, od dołu (za) kości potylicznej, a po bokach (poniżej) kości skroniowych.
Żylne zatoki mózgu
Ten post będzie poświęcony kilku małym mózgom o różnych strukturach mózgu niż ludzie, ale wyjątkowych zwiadowców, gadów i pszczół. Istnieje wiele innych możliwych przykładów bardzo małych zwierząt z inteligencją, takich jak mrówki i termity, ale wśród owadów najwięcej badań poświęcono pszczołom. Raport kończy się wskazówkami na temat tego, jak takie małe mózgi mogą mieć zaawansowane funkcje ze strukturami mózgu innymi niż ludzie.
Ptaki potrzebują stosunkowo dużego mózgu do latania. Ale w porównaniu z ludźmi i innymi dużymi zwierzętami ich mózgi są bardzo małe. Niedawno zauważono, że kakadu mają niezwykłą zdolność do korzystania z metod wieloetapowych w celu odblokowania bardzo złożonej komórki. Aby usunąć nakrętkę, ptak najpierw usunął kołek, następnie wykręcił śrubę, następnie wyjął śrubę, a następnie obrócił koło o 90 stopni, a następnie popchnął zatrzask na bok. Ptaki te były nieprzygotowane i były w stanie dowiedzieć się, jak to zrobić w mniej niż dwie godziny.
U owiec, w przeciwieństwie do bydła, część kości ciemieniowych bierze udział w tworzeniu tylnej części dachu jamy czaszki i, kierując się do okolicy potylicznej, tworzy grzebień kości ciemieniowej.
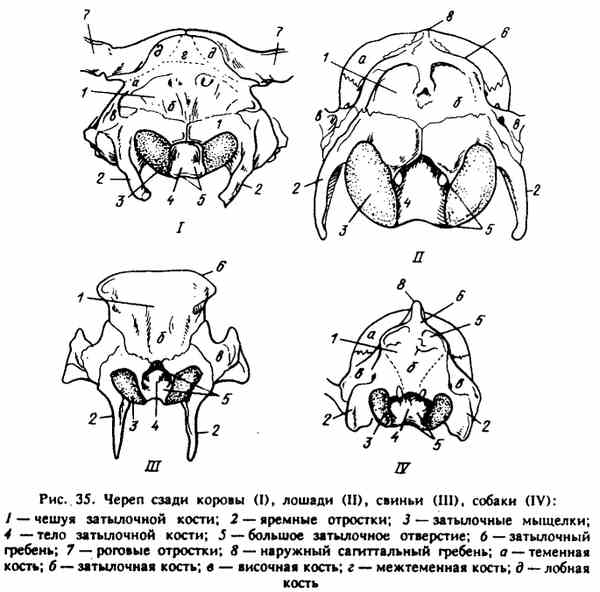
Ryc. 35. Czaszka za krową (I), koniem (II), świnią (III), psem (IV)
Tętnice mózgowe
Inni mogli się uczyć, obserwując. Ponadto, gdy tylko mogli otworzyć jeden z zamków, nigdy nie zapomnieli, jak mogą to zrobić natychmiast. Ponadto kakadu został ostatnio całkowicie obalony przez pogląd, że zwierzęta nie mogą tańczyć, a ponadto zauważają stałość obiektu na poziomie czteroletniej osoby lub małpy.
Jace, wraz z papugami, ptakami śpiewającymi i krukami, to jedne z najmądrzejszych ptaków. Ostatnio doniesiono, że podczas stada zmarłego towarzysza obserwowano całe stado sójek przez 48 godzin bez karmienia. Kaledoński kruk wykazał się wybitną pamięcią, w tym wspomnieniem ludzi i samochodów, które zirytowały ich po latach. Wykazują metapoznanie i liczenie, a także tworzą i używają zaawansowanych narzędzi lepiej niż wiele naczelnych. Zmodernizowali narzędzia, uformowali kawałek drutu w hak, aby uzyskać trudno dostępną przyjemność.
Kości czaszki charakteryzują się tym, że wewnątrz jamy czaszki kości mają gładką powierzchnię, na której widoczne są małe wgniecenia - wgniecenia, tak zwane odciski palców - odciskają cyfry. To są odciski mózgu.
TYLNA ŚCIANA JASKOŚCI JEDNORODNEJ (ryc. 35). Tworzy kość potyliczną - os occipitale. Ma trzy ogniska kostnienia enchondralnego, dlatego u młodych zwierząt dzieli się na trzy części: łuski - squamaoccipitalis, kłykciny (boczne) części - partes laterales i ciało kości potylicznej (część główna) - pars basilaris. Części boczne (kłykcina) otaczają duży otwór - otwór przedni dla wyjścia rdzenia kręgowego do kanału kręgowego. Przy silnym przechyleniu głowy w dół między kością potyliczną a atlasem tworzy się przestrzeń - dostęp do rdzenia kręgowego, który jest wykorzystywany podczas uboju zwierząt. Po bokach dużego otworu znajduje się zaokrąglony kłykc kości potylicznej - kłykcie potyliczne, które są pokryte chrząstką stawową, która wraz ze stawami stawowymi atlasu tworzy staw atlantokokowy.
Obok kłykcia przebiegają procesy szyjne - processus jugulares. Są bardzo potężne, proste u świń, nieco mniejsze u koni i przeżuwaczy oraz małe, lekko zaokrąglone u psów.
U podstawy procesów szyjnych po środku leży okrągły otwór gnykowy - otwór podniebienny, który wchodzi do kanału gnykowatego, przez który ostatnia - dwunasta para nerwów czaszkowych - główek - wyłania się z czaszki.
U dorosłego zwierzęcia wszystkie trzy części kości potylicznej są stopione. Łuski kości potylicznej u koni, świń i psów znajdują się powyżej kłykcia, a jej górna część tworzy potężny grzebień potyliczny (siedzący) - crista occipitalis nuche). Ma kształt trójkąta i u świń tworzy prostokąt.
DOLNA JASNOŚĆ JEDNORODZINNA (ryc. 36). Tworzy go główna część (ciało) kości potylicznej i kości sferoidalnej. Wzdłuż korpusu kości potylicznej powstaje poszarpana dziura - otwór przedni (czasami w postaci szczeliny u świń i koni), przez który wychodzą nerwy czaszkowe (gałęzie V, IX, X i XI).
Kość klinowa - os sphenoidale znajduje się przed podstawą czaszki. Jeśli izolujesz go od czaszki (w młodym wieku, dopóki kości nie stopią się w szwach), wygląda jak motyl. Środkowa / niesparowana część - ciało - ciałko sferyczne znajduje się przed ciałem kości potylicznej. Dwie pary cienkich płytek kostnych rozciągają się wzdłuż boków ciała. Przednie są nieco większe niż tylne - skrzydła kości sferycznej - alae sphenoidales, wraz z telomgonią tworzą przednią i środkową jamę czaszki, w której leży mózg pośredni i końcowy.
Zwróć uwagę na wewnętrzną powierzchnię ciała kości sferycznej, ma ona dwa etapy: górny przód jest prespenoidem, tylny i dolny są podstawowymi fenoidami, oddzielonymi od siebie wąską poprzeczną szczeliną, zwaną jelitem przecięcia wzrokowego - sulcus chiasmatis. U młodych zwierząt między nimi jest róża synchod. Na dolnym stopniu - podstawa fenyto, widoczny jest dołek, przechodzący z tyłu do wzniesienia (na granicy z ciałem kości potylicznej). Formacje te przypominają kształt azjatyckiego siodła, w związku z którym otrzymały nazwę „siodło tureckie” - sella turcica. W jego dolnej części znajduje się przysadka mózgowa (gruczoł dokrewny), przed którą w rynnie przecięcia wzrokowego znajduje się przecięcie nerwów wzrokowych, które przenoszą impuls z siatkówki do mózgu.
Na przedniej krawędzi zewnętrznej powierzchni skrzydeł kości sferycznej znajdują się otwory od góry do dołu, przez które nerwy czaszkowe łączą mózg z narządami głowy znajdującymi się z przodu ściany orbity. Najwyżej znajduje się otwór etoidalny - otwór przedni (jest również utworzony przez kość czołową), poniżej znajduje się układ wzrokowy canalis canalis opticum, pod którym koń i pies mają szczelinę oczodołową - fissura orbitalis, a nawet niższy okrągły otwór - otwór rotundum (u świń i przeżuwaczy połączone w otwór okrągłego oka).
Na zewnątrz kości kośćcowej znajdują się procesy pterygoidowekadrowanie hoansów. Są to duże i laminujące bydło i świnie. U koni i psów u podstawy tych procesów znajduje się kanał pterygoidalny - canalis pterygoideus, w którym przechodzą tętnica szczękowa i nerw.
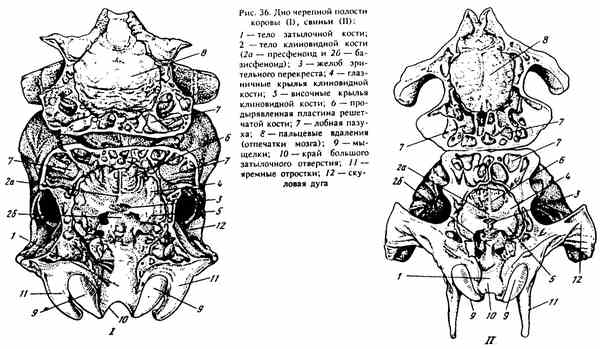
Ryc. 36. Dno jamy czaszki krowy (I), świni (II)
BOCZNE ŚCIANY JASKINI ŻURAWIA. Z góry prawe i lewe sparowane skrzydła kości sferycznej są połączone szwami ze łuskami kości skroniowej i czołowej i tworzą boczne ściany jamy czaszki. W młodym wieku istnieje tu nieskostniały obszar - fontanel w kształcie klina - sferoidalny.
Kość skroniowa - os temporale bierze udział w tworzeniu ścianki bocznej jamy czaszki. Składa się z czterech części: łuszcząca się część - pars squamosa - płyta, która tworzy ścianę boczną; część kamienista - pars petrosa - w części ogonowej jamy czaszki; część bębna - pars tympanica - na zewnętrznej stronie czaszki, wewnątrz której wyróżnia się wewnętrzna część bębna - pars endotympanica.
Proces jarzmowy kości skroniowej, processus zygomatics os temporale, rozgałęzia się bocznie z zewnętrznej powierzchni skroniowych łusek kości, która pochyla się do przodu i łączy z procesem skroniowym kości jarzmowej, tworząc z nią łuk jarzmowy - arcus zygomaticus. Na brzusznej powierzchni blaszkowatego procesu jarzmowego znajduje się powierzchnia stawowa zlokalizowana w płaszczyźnie segmentowej do połączenia z dolną szczęką.
W przedniej części łuski i skrzydła skroniowe kości sferoidalne tworzą ściany boczne po wewnętrznej stronie jamy czaszkowej, a na zewnątrz ściany orbity, na powierzchni której znajduje się otwór etoidalny, kanał wzrokowy, szczelina oczodołowa i okrągły otwór, przez który wychodzi pięć nerwów czaszkowych (pary II, III, IV, VI) i gałęzie pary V), w wyniku czego ta część dna orbity jest jednym z najbardziej nasyconych nerwów czaszkowych.
Ogonowa część skroniowych łusek kości pokrywa kamienistą klatkę piersiową - pars petrosa (ryc. 37), która bierze również udział w tworzeniu ściany bocznej jamy czaszki i leży przed i wyżej od procesów szyjnych. Ta część jest najbardziej „niezależna” w przypadku (jest stosunkowo łatwa do oddzielenia), u innych zwierząt jest bardzo mocno połączona z łuskami. Część skalista została zaprojektowana głównie z myślą o narządach słuchu i równowadze. Jest widoczny z jamy czaszki, płaski, gładki, nosi ślady zwojów mózgu. Wewnątrz znajduje się labirynt kostny - wnęki kostne o różnych kształtach, w których znajdują się narządy słuchu i równowagi. Z tego labiryntu dochodzą dwa małe pęknięcia na powierzchni kamienistej części - zewnętrzne otwory kanalików ślimaka i akwedukt przedsionka - apertura externa vestibuli i apertura externa aqueductus ślimak.
Należy o tym pamiętać, ponieważ to przez nich jama kostnego labiryntu ucha wewnętrznego komunikuje się z przestrzeniami międzypowłokowymi jamy czaszki; W związku z tym choroba narządów słuchu może prowadzić do choroby opon mózgowych - zapalenia opon mózgowych.
Po stronie jamy czaszki w środkowej części powierzchni części kamienistej znajduje się dość duże zagłębienie, z którego prowadzą do kości dwa otwory: wewnętrzny słuchowy mięsień - meatus acusticus internus, przez który organ słuchu i równowagi jest połączony nerwem (VIII) z rdzeniem przedłużonym; otwór na nerw twarzowy (VII), który łączy mózg z mięśniami twarzy.
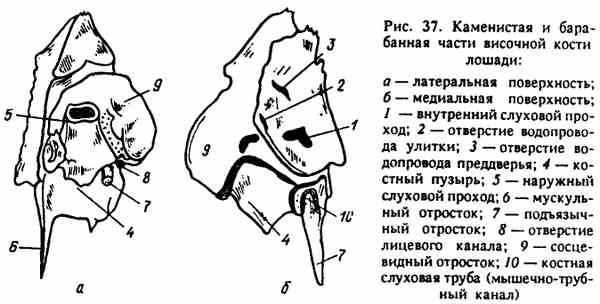
Ryc. 37. Kamieniste i bębenkowe części kości skroniowej konia
Na zewnętrznej stronie kamienistej części czaszki część bębna, pars tympanica, znajduje się poniżej, a proces wyrostka sutkowatego, processus mastoideus, znajduje się powyżej. Na części bębna wyraźnie widoczna jest znacząca (szczególnie u przeżuwaczy i świń) bańka bębnowa - bulla tympanica. Wewnątrz znajduje się jama - cavum tympani (lokalizacja ucha środkowego), do którego prowadzi górna rura kostna o bardzo dużej średnicy. Ta rurka jest zewnętrznym ujściem słuchowym - meatus acusticus externus, przewodzącym dźwiękiem, który przechodzi od ucha zewnętrznego przez środek do wnętrza. Poniżej, u podstawy zewnętrznego przewodu słuchowego, wyraźnie widoczny jest proces hoidalny (styloidalny) - processus hyoideus (stylordeus), który charakteryzuje się wyglądem zaostrzonego cylindrycznego kostnego patyka (szczególnie u konia). Kość gnykowa -os hyoideum jest przyczepiona do procesu gnyka, na którym zawieszony jest język, gardło i krtań. Tutaj widać ostry proces mięśniowy, do którego są przyczepione mięśnie kurtyny podniebiennej. U podstawy widoczna jest duża szczelina, prowadząca również do jamy pęcherza bębenkowego, w której znajduje się ucho środkowe. Luka ta to kanał mięśniowo-jajowodowy (rurka słuchowa) - canalis muscu-lotubarius (tuba audiva), przez który ucho środkowe komunikuje się z jamą gardła. Jest to droga zakażenia od gardła do ucha środkowego.
Proces wyrostka sutkowego - pocessus mastoideus jest dość obszerny, zbudowany z gąbczasta substancjapokryte cienką warstwą wyprasek. Do niego należy zastosowanie aparatu słuchowego w przypadku wykluczenia możliwości przeprowadzenia fali dźwiękowej przez ucho środkowe.
Pod procesem wyrostka sutkowatego znajduje się otwór szyjno-wyrostka sutkowatego - otwór czołowy Jomastoideum, przez który nerw twarzowy (VII) opuszcza część kamienistą w celu unerwienia mięśni twarzy.
Podstawa kamienistej części na granicy z ciałem kości potylicznej może uczestniczyć w tworzeniu poszarpanej dziury.
PRZEDNIE ŚCIANKI JASKINI KRÓLEWSKIEJ. Przed kością sferoidalną, tworzącą przednią ścianę jamy czaszkowej, znajduje się kość sitowa - os ethmoidale. Ta kość nie jest widoczna na powierzchni czaszki. Leży na granicy między jamą czaszki i nosa. Można go zobaczyć od wewnętrznej strony przedniej ściany jamy czaszki w postaci dwóch zaokrąglonych wgłębień, na dole których widoczne są płytki z masą otworów, jest to płyta perforowana - blaszka cribrosa. W otworach tej płytki nerwy przechodzą do żarówek węchowego mózgu, łącząc mózg z narządem węchowym jamy nosowej.
Od strony jamy nosowej do perforowanej płytki przymocowana jest masa najcieńszych płytek kostnych tworzących loki o różnych rozmiarach. Te zewnętrzne i wewnętrzne małżowiny są endoturbinal i ektoturbinal. W sumie te loki tworzą labirynt - labitynty ethmoidalis. Prawa i lewa część labiryntu są oddzielone dość cienką płytką umieszczoną wzdłuż środkowej linii strzałkowej, która nazywa się płytką prostopadłą. Z boku jamy czaszkowej jej początek jest widoczny w postaci pionowo ustawionego przegrzebka umieszczonego między dwoma wgłębieniami perforowanej płyty.
Od strony jamy nosowej prostopadła płytka kości sitowej przebiega do przodu w postaci już chrzęstnej, nigdy nie kostniejącej przegrody nosowej. Od prostopadłej płytki od strony jamy nosowej po prawej i lewej stronie są cienkie faliste kostne płytki otaczające labirynt po bokach. Okazuje się, że główna część, kość sitowa, labirynt, w którym znajduje się narząd węchowy, leży pomiędzy trzema płytkami, które tworzą rodzaj „pudełka” z dwoma przedziałami (pudełko to jest podzielone na dwie symetryczne przedziały prostopadłą płytą). Tylna ściana pudełka (płyta perforowana) jest widoczna z boku jamy czaszki. Od strony jamy nosowej przymocowany jest do niej labirynt. To pudełko jest otwarte tylko od strony jamy nosowej.
Aby lepiej zrozumieć lokalizację i nazwę wszystkich otworów, przez które mózg leżący w jamie czaszki komunikuje się z narządami, do których przechodzą jego nerwy, spójrz ponownie na te otwory na czaszce. Na przedniej ścianie, przez otwory perforowanej płytki kości sitowej, jama czaszki łączy się z labiryntem kości sitowej, tj. Z jamą nosową. Na tylnej ścianie, przez duży otwór (potyliczny), mózg przechodzi do rdzenia kręgowego, a jama czaszki komunikuje się z kanałem kręgowym. Po bokach podstawy czaszki utworzonej przez ciała kości potylicznej i kości sferycznej znajduje się szereg otworów, przez które nerwy czaszkowe łączą mózg z różnymi narządami głowy i ciała. Na zewnętrznej powierzchni przedniej części ściany orbity od góry do dołu przebiega łańcuch pionowo rozmieszczonych otworów: etoidalny, kanał wzrokowy, szczelina oczodołowa oraz okrągły, rozdarty otwór nieco ogonowy wzdłuż korpusu kości potylicznej, za którym znajduje się okrągły podjęzykowy otwór z boku przyśrodkowej powierzchni podstawy szyjnej ( podjęzykowy kanał nerwowy).
Muszle mózgowe
Żylne zatoki mózgu
Tętnice mózgowe
Ogólna charakterystyka struktury mózgu
Ogólna charakterystyka rozwoju mózgu
Koniec mózgu - telencefalon
Rozwój mózgu
Diencephalon-diencephalon
Diencephalon
Śródmózgowo-śródmózgowia
Rozwój śródmózgowia
Diamentowy rombencefalon
Rozwój diamentów
Lista wykorzystanej literatury:
Muszle mózgowe
Dura mater - dura mater encephali - najbardziej zewnętrzny. W przeciwieństwie do twardej skorupy rdzenia kręgowego rośnie wraz z okostną kości czaszki, więc nie ma miejsca na zewnątrzoponowe. Między okostną a oponą twardą przechodzą tylko żyły, tworząc dwa układy zatok żylnych - grzbietowy i brzuszny.
Dwie fałdy rozciągają się do przestrzeni podtwardówkowej opony twardej: półksiężyc i błoniasty kontur móżdżku. Sierp składający się z falx cerebri- biegnie w kierunku strzałkowym od grzebienia koguta kości sitowej do móżdżku. Leży w podłużnej szczelinie między półkulami mózgowymi. Najbardziej sierpowaty fałd wyraża się u psa i konia - prawie osiąga ciało modzelowate; jest mniej u świń i krów, a zwłaszcza u małych przeżuwaczy. Namiotowy namiot móżdżkowy - tentorium cerebella membranaceum - odchodzi od śladu kostnego lub kości ciemieniowej, a także od grzbietów kamieniste kości i znajduje się w poprzecznej szczelinie między dużym mózgiem a móżdżkiem.
Pajęczynówka błony mózgowo-pajęczej encefali - na zwojach mózgu mocno połączonych z miękką skorupą. Przestrzeń podpajęczynówkowa jest zachowana tylko w szczelinach i bruzdach między zwojami i na podstawowej powierzchni mózgu, gdzie tworzy rozszerzenia - brzuszne spłuczki rdzenia przedłużonego, most cysterny, robak (i \u200b\u200binne). Pod błoną pajęczynówkową wzdłuż fałdy półksiężyca kosmki wychodzą w przestrzeń podtwardówkową, czasami przyjmując postać guzków - granulki pachyon - granulacje arachnoidalne. Granulki Pachyon wprowadza się do zatoki strzałkowej i zwiększają odpływ płynu mózgowo-rdzeniowego do żył.
Miękki lub naczyniówki, pia mater encephali - bardzo mocno połączony z mózgiem, wchodzi we wszystkie pęknięcia i wnęki i wraz z naczyniami przenika do substancji mózgu. Wracając z powrotem do jamy mózgu, bierze udział w tworzeniu opon naczyniowych-tela chorioidea, w których osadzony jest splot naczyniowy-splot naczyniówkowo-naczyniowy. Takie formacje znajdują się w komorach mózgu: boczny, trzeci i czwarty.
Płyn mózgowo-rdzeniowy płynu cerebrospinalis - przestrzenie podtwardówkowe i podpajęczynówkowe mózgu i rdzenia kręgowego są wypełnione i przez sparowane otwory - apertura lateralis ventricularis quartiududal z bocznych nóg móżdżku - a nie sparowany otwór - apertura mediana ventriculi quarti - komunikuje się z jamami komór i kanałów mózgowych. Płyn mózgowo-rdzeniowy jest wydzielany przez wyściółkę i komórki splotu naczyniowego. Płynie w przestrzeni podpajęczynówkowej w kierunku mózgu, czyli czaszkowo i ogonowo w centralnym kanale rdzenia kręgowego. Odpływ płynu mózgowo-rdzeniowego następuje z przestrzeni podtwardówkowej do układu żylnego, a z przestrzeni podpajęczynówkowej do narządów limfatycznych.
Żylne zatoki mózgu
Układ zatoki grzbietowej składa się z niesparowanej strzałkowej i bezpośredniej zatoki oraz sparowanego pieprzu, kamieni potylicznych i grzbietowych; płyną do nich żyły mózgu.
Strzelec zatokowy strzałkowy znajduje się w fałdzie półksiężyca; nosowo, zaczyna się od żył błoniastych, u konia także od żył sitowych i mózgowych. Wpadają w to:
1) żyły grzbietowe mózgu-vv.cerebri dorsales;
2) błoniaste żyły opony twardej;
3) żyły kostne-vv. diploiki;
4) krótka prosta zatokowa zatokowa prosta.
Ten ostatni powstaje ogonowo z ciałka modzelowatego, przez połączenie żył ciałka modzelowatego z dużą żyłą mózgu.
Ciało modzelowate vein-v. corporis callosi - przenosi krew z przedniej części mózgu.
Wielka żyła mózgu-v. cerebri magna - bierze w głębokie żyły mózgu-vv. cerebri profundi przenoszący krew z mózgu i splotów naczyniowych.
Ogonowo od zbiegu zatoki bezpośredniej zatok strzałkowy dzieli się na prawe i lewe zatoki poprzeczne, -sinus transversus - z których każdy jest skierowany na kanał skroniowy i przechodzi do grzbietowej żyły mózgowej-v. cerebri dorsalis. Ten ostatni wpływa do żyły powierzchownej skroniowej. Kamienista zatokowa petrosus-zatokowa, która przenosi krew z podstawowych części mózgu, wpada do zatoki poprzecznej, zanim wpada do kanału skroniowego.
Pomiędzy zatokami poprzecznymi znajduje się łączna komunikacja zatok-zatok - leży ona u podstawy namiotu móżdżku. Zatoki potyliczne wpadają do niej zatoki potyliczne, znajdujące się w bruzdach po obu stronach robaka móżdżku.
Odpływ krwi z układu grzbietowego zatok, oprócz mózgowych żył grzbietowych, występuje również przez emissarium emissarium do żył głębokich skroniowych. Wysłannicy to nietrwałe kanały wyładowcze perforujące ścianę jamy czaszki.
Brzuszny lub podstawowy układ zatok obejmuje sinusy okrągłe i podstawowe.
Okrągła zatokowa zatokowa okrągła przysadka mózgowa. Tworzą ją prawe i lewe zatoki jamiste, łączące między jamiste zatoki-zatoki cavernosus et intercavernosus.
Każdy zatok jamistych przechodzi nosowo do żyły oczodołowej mózgu ogonowo do zatoki podstawnej. Żyła orbitalna mózgu-v. cerebralis orbitalis - przepływa przez szczelinę oczodołową do głębokiej żyły twarzy.
Zatoka podstawna zatoki podstawnej - brzuszna zatokowa część kręgosłupa. W obszarze poszarpanej dziury daje żyłę brzuszną i mózgową-v. cerebralis ventralis - który wpływa do żyły potylicznej. W pobliżu stawu potylicznego zatoki podstawne zespalające się między sobą tworzą brzuszną zatokę potyliczną potylicy potylicznej. Brzuszne żyły mózgu otwierają się na podstawowy system zatok.
Tętnice mózgowe
1. Wewnętrzna tętnica szyjna-a. carotis interna - Wnika do jamy czaszki przez otwór szyjny lub rozdarty i dzieli się na gałęzie łączące nos i ogon. Z gałęziami o tej samej nazwie po drugiej stronie, tworzą się one w tętnicy przysadkowej krążenia tętniczego móżdżku. Tętnica nosowa-a wychodzi z pierścienia. cerebri nasalis - trafia do ciała modzelowatego i karmi przednie półkule.
Odgałęzienie 4 naczyń z gałęzi nosa:
a) tętnica nosowa opon mózgowych-a. meningea nasalis;
b) środkowa tętnica mózgowa -a. media cerebri trafiają na boczną powierzchnię mózgu;
c) tętnica nosowa splotu naczyniowego-a. chorioidea nasalis- wzdłuż drogi wzrokowej podąża za splotem naczyniowym komory bocznej;
d) wewnętrzna tętnica orbitalna -a. okulistyka międzynaro- dowa przechodzi przez nerw wzrokowy na orbitę.
Z gałęzi łączącej ogonowo:
a) ogonową tętnicę mózgową -a. cerebri caudalis - na tylne półkule i poczwórne
b) tętnica ogonowa splotu naczyniowego-a. chorioidea caudalis - w splocie naczyniówkowym komór bocznych.
2. Z tętnicy potylicznej tętnica kręgowa - a odchodzi. cerebrospinalis - przez międzykręgowy otwór atlasu przenika kanał kręgowy i dzieli się na gałęzie czaszki i ogona, łącząc się z tymi samymi gałęziami po drugiej stronie. Główna tętnica mózgu-a jest oddzielona od gałęzi czaszki w mózgu. basilaris cerebri - wpada do pierścienia tętniczego, a po drodze daje: za mostem jest tętnica ogonowa móżdżku, przed mostem jest tętnica czaszkowa móżdżku-a. cerebella caudalis et cranialis - i do nerwu słuchowego - tętnica słuchowa wewnętrzna - a. audiva interna. Ogonowa gałąź zespoleń rdzenia kręgowego z rdzeniem tętnicy brzusznej -a. spinalis ventralis. Naczynia mózgu są unerwione przez współczulny układ nerwowy.
Ogólna charakterystyka struktury mózgu
Mózg - mózg - z powierzchni grzbietowej jest podzielony poprzeczną szczeliną - fissura transversa cerebri - do dużego i romboidalnego mózgu.
Mózg mózgowy składa się z dwóch półkul, móżdżku i śródmózgowia. Prawa i lewa półkula mózgu - hemisphaerum cerebri dextrum et sinistrum - są podzielone od strony grzbietu głęboką szczeliną podłużną, fissura longitudinalis cerebri. Pośredni i śródmózgowie są pokryte grzbietowo półkulami.
Mózg romboidfalno-romboidalny składa się z móżdżku, mostka mózgowego połączonego z tylną częścią mózgu i rdzenia przedłużonego.
Móżdżek móżdżek znajduje się grzbietowo od rdzenia przedłużonego i tylnej części półkul mózgowych.
Medulla oblongata-medulla jest bezpośrednią kontynuacją rdzenia kręgowego. Z przodu znajduje się most cerebri-pons cerebri.
Przed mostem znajdują się nogi dużego mostu. Z przodu nogi rozciągają się na drogi wzrokowe - tractus opticus - i wzrokowy chiasm-chiasma opticum. Pomiędzy nogami dużego mózgu, za skrzyżowaniem wzrokowym, znajdują się: wyrostek sutkowy-ciało mamillare- i szary guz z lejkiem i przysadką mózgową. Przed drogami wzrokowymi i po bokach nóg mózgu mózgowy węchowy rhinencephalon wystaje; doustnie kończy się dwiema cebulkami węchowymi, bulbi olfactorii. Mózg węchowy reprezentuje brzuszną część półkul mózgowych oraz część peleryno-palio-grzbietowo-boczną pokrytą bruzdami i zwojami.
Półkule należą do terminalnego mózgu-telencefalonu - a dróg wzrokowych, szarego guzka, lejka, przysadki i ciała sutkowatego - do brzusznych części diencefalonu diencefalonu.
Nogi dużego mózgu to brzuszna część śródmózgowia mezenecefalonu.
Móżdżek i mostek są częścią śródmózgowia-mózgu. Pośredni, rdzeń i śródmózgowia są połączone w pień mózgu - pień mózgu - cerebri.
Ogólna charakterystyka rozwoju mózgu
Mózg rozwija się w związku z pojawieniem się i dalszym rozwojem różnych wyższych ośrodki nerwowe: receptor, motoryczny i asocjacyjny, które zapewniają skoordynowaną funkcję wszystkich narządów - somatycznej, trzewnej, naczyniowej.
We wczesnych stadiach ontogenezy i filogenezy mózg jest powiększonym końcem rurki mózgowej, która leży przed cięciwą, dlatego nazywa się ją przedchordowym lub pierwotnym pęcherzem mózgowym. Jego rozwój związany jest z narządem węchu, a jego funkcja węchowa jest zachowana u wszystkich zwierząt, w tym ssaków.
Nieco później, na przednim końcu mózgu epichordalnego, izolowany jest przedchordowy lub wtórny pęcherz mózgowy. Jego rozwój wynika z: pojawienia się pierwotnego centrum stowarzyszeniowego i komisyjnego; obecność aparatu skrzelowego i narządów linii bocznej zwierząt wodnych; różnicowanie narządów wewnętrznych.
Z przedniej części epichordalnego pęcherza mózgowego izoluje się środkowy pęcherz mózgowy, który, z wyjątkiem ssaków, odgrywa wiodącą rolę, ponieważ impulsy pochodzą zdecydowanie ze wszystkich aparatów receptorowych, a zwłaszcza wzrokowych.
Od powierzchni grzbietowej wszystkie trzy części mózgu są dość ostro oddzielone od siebie poprzecznymi wargami włókien nerwowych, z przodu i za śródmózgiem.
Następnie mózg prechordalny dzieli się na mózg terminalny w postaci sparowanego pęcherza i diencefalonu.
Mózg końcowy osiąga najwyższy etap rozwoju u ssaków z obecnością nowej kory mózgowej, która staje się wyższym ośrodkiem nerwowym.
W diencephalon u niższych zwierząt, w związku z wiodącą rolą analizatora węchowego, pierwsze miejsce zajmują podwzgórze i nabłonek. Są to pośrednie centra węchowe, które wysyłają impulsy do śródmózgowia, a podwzgórze dodatkowo wysyła impulsy do przysadki mózgowej i do autonomicznych centrów środkowego i rdzeniowego przedłużenia. Z tego powodu podwzgórze staje się najwyższym podkorowym ośrodkiem wegetatywnym. Chroni również tę funkcję u ssaków.
Tylko wraz z tworzeniem kory mózgowej u ssaków środkowa część diencefalonu, wzgórze, przez które kora decydująco odbiera wszystkie impulsy receptora z peryferii, zaczyna intensywnie się rozwijać.
Tak więc historycznie podwzgórze jest bardziej związane z mózgiem epichordalnym, a wzgórze z przedgordowym.
Niemal równocześnie z podziałem mózgu prechordalnego mózg epichordalny różnicuje się za śródmózgiem w śródmózgowie i rdzeń przedłużony. Mózg ssaków jest reprezentowany tylko przez móżdżek, który jest podkorowym centrum korelacji ruchów mięśni w celu utrzymania równowagi. Tylko u ssaków mostek mózgowy jest dodawany do móżdżku ze względu na rozwój nowej kory mózgowej, a sam móżdżek jest wzmacniany przez dwie półkule mózgowe.
Wnęki pierwotnych pęcherzy mózgowych stają się komorami mózgowymi w rozwiniętym mózgu. Z jamy mózgu przedskordowego w mózgu terminalnym powstają sparowane komory boczne, aw diencefalonie trzecia komora mózgowa. Wszystkie trzy komory są połączone otworem międzykomorowym. Komora środkowego pęcherza mózgowego z powodu wzrostu ścian ostatniej transformacji do akweduktu mózgowego, a romboidalna jama staje się czwartą komorą mózgową, która komunikuje się z centralnym kanałem kręgowym.
Koniec mózgu - telencefalon
Składa się z dwóch półkul mózgowych - hemisphaerum dextrum et sinistrum - oddzielonych od powierzchni grzbietowej głęboką szczeliną podłużną - fissura longitudinalis cerebri.
Na każdej półkuli badany jest płaszcz, mózg węchowy, prążkowia i boczne komory mózgu. Płaszcz znajduje się na półkuli grzbietowo od komory, mózg węchowy jest wentylacyjno-przyśrodkowy. Granica między płaszczem a mózgiem węchowym na brzusznej powierzchni mózgu jest podstawowym rowkiem granicznym. Prążkowia leży w brzusznej ścianie półkuli grzbietowo od części mózgu węchowego.
A) Płaszcz-pal
Składa się z szarej i białej materii mózgowej. Szary rdzeń rdzeniowy tworzy na swojej powierzchni korę mózgową - kora mózgowa -. Na nim rozróżnij zwoje wirusa mózgu - oddzielone bruzdami i szczelinami. Trzy łukowate wiatraki są widoczne na bocznej powierzchni płaszcza. Otaczają boczny sylvian sulcus-sulcus lateralis. Łukowate zwoje są wyraźnie wyraźne u psa, au innych zwierząt każde łukowate jest podzielone na części przez bruzdy. Na środkowej powierzchni peleryny widoczne są głównie dwa łukowate wiatraki wokół ciała modzelowatego, a najbliższy zakręt zakrętu obręczy, funkcjonalnie połączony z mózgiem węchowym.
Biały rdzeń płaszcza alba znajduje się pod korą płaszcza. Składa się ze ścieżek - asocjacyjnej, komisjonalnej i projekcji.
a) włókna asocjacyjne łączą oddzielne sekcje kory w obrębie każdej półkuli. Są one podzielone na krótkie i długie włókna.
b) włókna komisuralne łączą obszary należące do różnych półkul. Tworzą one corpus callosum corpus corpus, największe pęknięcie mózgu. Jest on umieszczony między półkulami na głębokości szczeliny podłużnej. Rozróżnij tułów korpusu callosi truncus corporis corporis i dwa końce - przód i tył. Przedni koniec nazywa się kolanem ciałka modzelowatego - genu corporis callosi - wygina się brzusznie. Tylny koniec lub grzbiet korpusu callosi-splenium corporis corporis łączy się z łukiem.
Włókna komorowe wyłaniające się z tułowia modzelowatego tworzą blask komisyjnej radiatio corporis callosi - tworzy ścianę grzbietową bocznej komory mózgu i rozchodzi się w przednią, boczną i tylną część kory peleryny.
c) włókna projekcyjne - połącz korę płaszcza, zarówno z poszczególnymi częściami pnia mózgu, jak i za pomocą rdzeń kręgowy. Tworzą wewnętrzną kapsułkę w prążkowiu. Funkcjonalnie ścieżki projekcji są podzielone na eferentne i aferentne.
Ścieżki przepływowe przenoszą impulsy z kory półkul mózgowych do różnych części pnia mózgu: dużego, romboidalnego i rdzeniowego.
Ścieżki aferentne dostarczają impulsy do kory półkul z rdzenia kręgowego i mózgu w kształcie rombu. Tworzą one tył wewnętrznej kapsułki.
W przeciwieństwie do kory mózgowej, cała szara substancja pozostałych części całego ośrodka układ nerwowy zjednoczona przez koncepcję „podkorty”. Impulsy ze wszystkich części ciała najpierw trafiają do różnych części kory mózgowej, w tym do guzka wzrokowego, a z tego drugiego wchodzą do kory półkul. Kora stale się rozkłada, dezintegruje wzbudzenia powstające w urządzeniach receptorowych, a jednocześnie łączy, syntetyzuje działanie wzbudzeń izolowanych przez analizę na różne kompleksy. Jest to przejaw analitycznej i syntetycznej aktywności kory mózgowej. Co więcej, najwyższa część układu nerwowego - kora mózgowa (u ludzi) jest materialnym substratem wyższej aktywności nerwowej: myślenia, świadomości, pamięci, rozumu, czyli wszelkiej aktywności umysłowej, która jest wynikiem fizjologicznej aktywności kory mózgowej.
B) węchowy mózg-rhinencephalon
Znajduje się on w części wentylacyjno-przyśrodkowej każdej półkuli dużego mózgu. Niektóre jego części są widoczne na podstawowych i środkowych powierzchniach półkul, a także na dnie bocznych komór mózgu. Cebule węchowe, drogi węchowe i zwoje, trójkąty węchowe i płaty w kształcie gruszki znajdują się na podstawowej powierzchni półkul. Na środkowych powierzchniach półkul, pole węchowe, zwoje hipokampa, zwoje obręczy i powierzchnia nacięcia zrostów nosowych są widoczne, a jądro ogoniaste, hipokamp i łuk są widoczne u dołu bocznych komór mózgu. Zakręt lędźwiowy, hipokamp, \u200b\u200błuk i ciało migdałowate są układem limbicznym - pars limbica rhinencephalon.
1. Cebula-bulbus olfactorius - sparowane formowanie w postaci dość płaskiego, wydłużonego i zakrzywionego grzbietowego procesu mózgowego, który wystaje poza przednią krawędź półkuli mózgu w węchową dziurę kości sitowej. Część grzbietowo-żarowa żarówki jest zbudowana z szarej rdzenia, a boczno - brzuszna - z białej rdzenia. W opuszce znajduje się komora opuszki węchowej - ventriculus bulbi olfactorii - jest ona kontynuacją bocznej komory mózgu. Opuszka węchowa obejmuje nerw węchowy-n. olfactorius. Zawiera liczne wiązki włókien nerwowych - fila olfactoria - skierowane z komórek węchowych błony śluzowej nosa do komórek nerwowych opuszki. Zatem żarówki węchowe są głównymi ośrodkami węchowymi.
2. Od komórki nerwowe żarówka węchowa rozpoczynają się szlaki węchowe . Tworzą one biały rdzeń samej cebulki i drogi węchowe - pospolity, przyśrodkowy i boczny pąk olfactorius communis, medialis i lateralis. Boczny przewód węchowy przechodzi do płata w kształcie gruszki, pokrywając boczny zakręt węchowy girus olfactorius lateralis na całej swojej ścieżce. Przyśrodkowy przewód węchowy dociera do środkowej powierzchni peleryny, tworząc para-węchową parolfaktorię polową. Układy węchowe ograniczają trójkąt węchowy, trygon olfactorium, z istoty szarej. Przewodzą impulsy z opuszki węchowej do komórek wtórnych ośrodków węchowych w zakręcie węchowym, trójkątach węchowych, polach węchowych, w płatach w kształcie gruszki, a także w podwzgórzu i śródmózgowiu.
3. Lobus periformis w kształcie gruszki - położony przyśrodkowo od bocznego przewodu węchowego i ogonowo od trójkąta węchowego; środkowo graniczy z nogami dużego mózgu. Ogonowo-przyśrodkowa krawędź płatka w kształcie gruszki jest przyśrodkową szczeliną graniczną lub hipokampami Fissura. W płatku w kształcie gruszki znajduje się wnęka reprezentująca tylną część bocznej komory mózgu.
Płat w kształcie gruszki ogonowo przechodzi bez wyraźnej granicy do zakrętu hipokampa znajdującego się na środkowej powierzchni półkuli, z tyłu i z boku od szczeliny hipokampa.
Zakręt hipokampa grzbietowo przechodzi do obręczy zakrętu wirowego zakrętu wirowego. Ten ostatni przechodzi bezpośrednio od strony grzbietowej modzelowatego i zginając się wokół niego, łączy się z polem węchowym i jest częścią kory płaszcza. Płat w kształcie gruszki jest wtórnym centrum węchowym.
4. Hipokamp lub róg hipokampa Ammon - z częścią grzbietową tworzy dno bocznej komory mózgu, za jądrem ogoniastym, od którego oddziela się splot naczyniowy komory bocznej. Hipokamp jest fałdem kory mózgowej w obszarze szczeliny hipokampa i płata piriformalnego. Półksiężyc wygina się bocznie 0-ogonowo i brzusznie i gubi się w ścianie płata w kształcie gruszki. Rogi amonowe leżą grzbietowo na guzkach wzrokowych, oddzielone od nich splotem naczyniowym trzeciej komory mózgowej. Jako najwyższe asocjacyjne podkorowe centra węchowe i smakowe rogi amonowe są związane z różnymi częściami kory mózgowej i jąder podkorowych. Ich ścieżki przewodzące tworzą łuk i jego pochodne.
5. Vault -fornix - zawiera ścieżki łączące hipokamp z ciałem sutkowatym diencephalonu. Oddzielne sekcje tego pakietu włókien tworzą ryflowany arkusz, brzeg rogów amonowych, nogi, kolumny, ciało i przednią komisję mózgu.
Rowkowany hipokamp liściowo-pęcherzykowy pokrywa hipokamp swoją powierzchnią skierowaną w stronę bocznej komory mózgu. Tworzą go włókna nerwowe pochodzące z szarego rdzenia płata w kształcie gruszki i hipokampa. Na marginesie grzbietowo-bocznym włókna te tworzą granicę hipokampa hipokampa-fimbrii rozciągającego się nosowo do nogi łucznika krzyżowego. Ta ostatnia, łącząc się z nogą, staje się z kolei krótkim ciałem łukowatego korpusu rozwidlenia. Ciało łuku służy jako ściana grzbietowa trzeciej komory mózgowej; nosowo jest podzielony na dwie kolumny arch-columnae fornicis. Te ostatnie przyśrodkowo z jąder ogoniastych są kierowane do ciała wyrostka sutkowego i szarego pagórka podwzgórza. Szczelina łuku - comissura fornicis - jest utworzona przez poprzeczne włókna między nogami łuku; łączy ze sobą grzbietowe końce rogów amonowych.
6. Pomiędzy przednim końcem ciała modzelowatego a filarami łuku znajduje się przednia komisura mózgu-komissura nasalis. Łączy mózg węchowy obu półkul ze sobą.
C) Striatum corpus striatum
Znajduje się przed wizualnymi guzkami, grzbietowo od trójkątów węchowych, składa się z czterech jąder: ogoniastego, soczewkowego, migdałowego i płotowego. Pomiędzy jądrami znajdują się wewnętrzne i zewnętrzne zrosty kapsułki utworzone przez ścieżki projekcji.
Na jądrze ogoniastym, jądrze ogoniastym wyróżnia się głowa i ogon. Głowa jądra ogoniastego z powierzchnią grzbietową tworzy dno bocznej komory, nosowo-bocznie od hipokampa.
Jądro soczewkowe, jądro lentiformis, znajduje się bocznie od głowy jądra ogoniastego i guzka wzrokowego i jest oddzielone wewnętrzną kapsułką-torebką wewnętrzną. Boczna część jądra soczewkowego nazywa się skorupą skorupy. Przyśrodkowa część soczewkowego jądra starszego pochodzenia nazywa się jasnym jądrem-globus pallidus. Jest połączony z mózgiem węchowym, leży bocznie od guzka wzrokowego i przed jądrem bocznie zakrzywionego zwoju ciała zwojowego. Zewnętrzna kapsułka-torebka zewnętrzna znajduje się bocznie od jądra soczewkowego, a drugie jądro, bocznie od kapsułki, jest drugim jądrem w postaci wąskiego paska miasta-klaustrum. Na granicy między płotem, skorupą i rogiem amonowym znajduje się jądro migdałowate, jądro amigdale, czyli mózg węchowy.
Ciała w paski są połączone ścieżkami: 1) z korą mózgową; 2) z wizualnymi guzkami i podwzgórzem; 3) z jądrami tworzenia siatkowej śródmózgowia, z jądrami mostka i rdzenia przedłużonego; 4) z jądrami nerwów czaszkowych.
Poprzez pasiaste ciała różne łańcuchy odruchowe są zamknięte lub:
A) aparat receptorów obwodowych - guzki wzrokowe - prążkowia - aparat efektorowy somatyczny i trzewny.
B) kora - prążkowie - somatyczny i trzewny aparat efektorowy.
U ssaków jądra prążkowia są najważniejszymi podkorowymi ośrodkami ruchowymi:
1) Skoordynowane ruchy mimowolne;
2) Regulacja napięcia mięśniowego w stanie spoczynku i ruchu;
3) Bezwarunkowe odruchy;
4) Wyższe podkorowe ośrodki wegetatywne
Ciała w paski działają jako całość, ale ich oddzielne części działają odwrotnie.
Rozwój mózgu
Ze starszych przednich i brzusznych ścian terminalnego mózgu pochodzą płaty węchowe i prążkowia, a od późniejszej ściany grzbietowej płaszcz.
1. Rozwój płatków węchowych mózgu jest determinowany obecnością analizatora węchowego, który u zwierząt wodnych ma szczególne znaczenie, gdy jest zorientowany w środowisku zewnętrznym. Procesy wrażliwych komórek węchowych kończą się w opuszkach węchowych, które tworzą część płatów węchowych mózgu końcowego, których proksymalne odcinki tworzą najstarszą korę węchową, palaeocortex, z szarej rdzenia. U ssaków tworzy zakręt węchowy i trójkąty węchowe - trigonum olfactorium - które są wtórnymi ośrodkami węchowymi.
2. W brzusznej ścianie końcowego mózgu, grzbietowo od kory węchowej, podstawa zwojowo-zwojowa podstawna pojawia się bardzo wcześnie. To najwyższe centrum motoryczne. Podstawowy zwoj zachowany jest u ssaków jako blade jądro. Później u zwierząt lądowych rosną dodatkowe jądra drobnokomórkowe, tworząc w całości - neostriatum - które składa się ze skorupy, a u ssaków jądro ogoniaste. Ponadto izolowane są nowe jądra - ogrodzenie i ciało migdałowate. Wraz z pojawieniem się wtórnego płaszcza złożony zwojach podstawy jest przebity przez wewnętrzne i zewnętrzne kapsułki ze ścieżek, które prowadzą do kory i z powrotem. Z tego powodu pojawił się nowy termin - ciało w paski.
3. W ewolucji peleryny obserwuje się dwie formacje o różnych funkcjach i strukturach, nie licząc błoniastej prymitywnej peleryny-pallium membranaceum - charakterystycznej dla zwierząt wodnych i składającej się z wyściółki. U zwierząt lądowych najpierw pojawia się kora o prostszej, trójwarstwowej strukturze komórkowej. Powstają z niego wyższe centra węchowe: płaty w kształcie gruszki, zakręt hipokampa i ich pochodne. Wszystkie te części łączy koncepcja pierwotnego płaszcza-arhipallium.
Podstawowy płaszcz charakteryzuje się umiejscowieniem szarego rdzenia pod białym rdzenia. Archipallium, podobnie jak paleopallium, najlepiej wyrażać u makrosmatycznych zwierząt - makrosmatów - z ostrym zmysłem węchu, w przeciwieństwie do mikrosmatycznych mikrosmatów - w których zmysł węchu jest mniej rozwinięty.
U ssaków na granicy między płatem węchowym a płaszczem pierwotnym pojawia się wtórny płaszcz z neopalu. Płaszcz wtórny charakteryzuje się rozmieszczeniem powierzchni szarego rdzenia w postaci kory nad białym rdzenia. Podstawy pojawiają się u gadów. W miarę wzrostu nowy płaszcz jest wypychany z Arichipalluim na środkową i brzuszną. W tym samym czasie w obszarze szczeliny hipokampa powstaje fałd kory w postaci hipokampu, a sam nowotwór zajmuje całą grzbietowo-boczną część półkuli i, rozprzestrzeniając się ogonowo, pokrywa środkowy, a nawet śródmózgowie z ich powierzchni grzbietowej i bocznej.
Wzrost wtórnego płaszcza jest spowodowany wprowadzeniem nowych włókien nerwu projekcyjnego z diencefalonu, które są przewodnikami różnych analizatorów - skórnych, wzrokowych, słuchowych, mięśniowych, oraz rozwoju nowych neuronów i ośrodków asocjacyjnych, w tym tworzenia siateczki.
Kora wtórnego płaszcza u szeregu zwierząt jest niezwykle skomplikowana w swojej strukturze, jego funkcje i struktura są bardzo zróżnicowane, a wymiary rosną. U dużych ssaków płaszcz zwykle nosi zakręt i bruzdy. Takie zwierzęta nazywane są gyrencephali. Meandry w serii zwierząt nie są takie same. W niektórych przypadkach poruszają się głównie łukami wokół poprzecznego rowka Sylviana; u naczelnych żyruzy tworzą dwa układy - czołowy i ciemieniowy. Oba systemy są oddzielone bruzdą Sylvian. W trzeciej grupie zwierząt bruzda leśna jest nieobecna, a bruzdy biegną z przodu mózgu, wzdłużnie i z tyłu poprzecznie. Dlatego homologacja zwojów między zwierzętami różnych rzędów jest niezwykle trudna, aw niektórych przypadkach może być niemożliwa.
U małych zwierząt zakręt jest na ogół nieobecny. Największa liczba zwojów występuje u słoni i wielorybów. W ontogenezie zakręt pojawia się również nie od razu, ale w określonej sekwencji.
Diencephalon-diencephalon
Znajduje się za prążkowiem końcowego mózgu i przed śródmózgiem, pokryty grzbietowo wyściółką naczyniową trzeciej komory mózgowej i hipokampa. Diencephalon składa się z trzech działów różnego pochodzenia, struktury i funkcji: nabłonka, wzgórza i podwzgórza. Nabłonek tworzy opona naczyniowa trzeciej komory mózgowej, szyszynki i sparowanego węzła wędzidełkowego. Wzgórze składa się z guzków wzrokowych, między którymi znajduje się trzecia komora mózgu w kształcie pierścienia. Podwzgórze składa się z wizualnego występu z płytką końcową, szarego guzka z lejkiem i przysadką mózgową oraz ciała sutkowatego. Wszystkie części podwzgórza są widoczne na podstawowej powierzchni mózgu, między nogami dużego mózgu, za przecięciem nerwów wzrokowych.
Wzgórze Guzki wzrokowe - optyka thalami - są najbardziej masywną częścią diencephalonu. Naso-bocznie łączą się z jądrem ogoniastym prążkowia; guzki wzrokowe zostaną oddzielone od tego ostatniego paskiem granicznym-pasmami końcowymi, od kwadrupola poprzecznym rowkiem, a od siebie dolną krawędzią guzków wzrokowych, pokrytą wyściółką naczyniową trzeciej komory. Guzki stanowią liczne skupiska jąder szarej rdzenia. Największy z nich:
1. nasienie jądra-jądro nasalis thalami - leży w grubości nosa tuberculum tuberculum nasalis thalami- w części nosowo-przyśrodkowej guzka wzrokowego. Jest to najstarszy i jest centrum przełączania ścieżek aferentnych na ścieżki refleksyjne.
2. jądro ogoniaste jądro ogoniaste ogoniaste - zamknięte w grubości odcinka ogonowo-bocznego guzka bocznego guzowatego tuberculum lateralis thalami - i składa się z pośrednich ośrodków wzrokowych i słuchowych. Od przecięcia nerwów wzrokowych i ośrodków słuchowych. Od przecięcia nerwów wzrokowych, chiasma opticum, na podstawowej powierzchni mózgu zaczynają się drogi wzrokowe tractus opticum. Każdy przewód zgina się bocznie wokół wzgórza i przechodzi do zgiętego ciała bocznego, corpus geniculatum lateralis, które jest zagubione w jądrze ogonowym ścieżek wzrokowych, które przechodzą do kory mózgowej. Między bocznym korbowym korpusem a czworokątem wystaje przyśrodkowe korbkowe ciało korpusu geniculatum medialis. Łączy wzgórza ogonowe czworokąta z jądrem ogonowym guzka wzrokowego i jest pośrednim ośrodkiem słuchowym w drodze do kory mózgowej. Oba korby są połączone w 7.
3. boczne jądro-jądro lateralis thalami - jest centrum do przełączania ścieżek przewodzących analizatora skóry i analizatora układu mięśniowo-szkieletowego na ścieżki prowadzące do kory mózgowej i prążkowia.
4. przyśrodkowe jądro-jądro medialis thalami - jest pośrednim centrum motorycznym dla ścieżek od kory do jądra układ pozapiramidowy: do czerwonego jądra, do jąder nerwów czaszkowych i rdzenia kręgowego.
5. tworzenie sieci - formatio reticularis - znajduje się między jądrami i jest z nimi związany.
Trzecia komora mózgu - ventriculus tertius - znajduje się między guzkami nerwu wzrokowego, ma okrągły kształt, gdy wrasta w nie masa pośrednia guzków nerwu wzrokowego massa intermedia thalami. W ścianach komory znajduje się środkowy szary rdzeń rdzeniowy grisea centralis - znajdują się w nim podkorowe ośrodki wegetatywne. Trzecia komora komunikuje się z akweduktem mózgowym śródmózgowia, a za nosową komisją mózgu - comissura nasalis - z bocznymi komorami mózgu przez przedsionkowo-komorowy otwór międzykomorowy.
Nabłonek . Na brzegach dołu guzków wzrokowych widoczne są paski mózgowe guzków wzrokowych, rozstępów rdzeniowych, a na nich sparowany węzeł wędzidełka zwojowego habenula. Węzeł wędzidełka przechodzi do wędzidełka lub smyczy-habenula, na której wzmocniony jest nasadowy nasadowy nasadowy gruszkowaty kształt. Szyszynka jest gruczołem wydzielania wewnętrznego leżącym w dole między guzkami wzrokowymi a czwórką. Zwoje wędzidełka służy jako pośrednie centrum ścieżek odruchowych między mózgiem, jądrem piątej pary i jądrem śródmiąższowym.
Wyściółka naczyniowa trzeciej komory mózgowej-tela chorioidea ventriculi tertii - utworzony przez fałd płytki nabłonkowej miękkiej błony mózgu i splotu naczyniowego. Płytki nabłonkowe opony są przymocowane wzdłuż krawędzi dołu guzków i łuku. Opona naczyniowa oddziela guzki wzrokowe od rogów amonowych i od łuku, przenika przez otwór międzykomorowy do bocznych komór mózgu w postaci splotów naczyniowych bocznych komór mózgu-splotu naczyniówkowo-naczyniowego chorioideus ventriculi lateralis. Opona naczyniowa tworzy zwis-recesus suprapinealis przed szyszynką i bezpośrednio za ciałkiem modzelowatym.
Podwzgórze - podwzgórze, bardziej pradawna część podwzgórza - tworzy ścianę trzeciej komory w kierunku brzusznym z guzków wzrokowych, jest najwyższym podkorowym ośrodkiem wegetatywnym, łączy się z guzkami wzrokowymi, z mózgiem węchowym i śródmózgiem. Podwzgórze zawiera centra regulacji: oddychanie, krążenie krwi, temperatura, metabolizm, funkcje seksualne. Jego przednia część jest reprezentowana przez szary guzek, lejek i przysadkę mózgową, a tylna część przez korpus wyrostka sutkowego i ściany trzeciej komory.
Podwzgórze szara bulwa cinereum - leży bezpośrednio za przecięciem nerwów wzrokowych, między nogami dużego mózgu. Pośrodku szarego wzgórza znajduje się lejek-zatoczka infundibuli. Sam lejek-infundibulum jest cienkościenny, do którego przymocowany jest przysadka mózgowa.
Przysadka mózgowa - jest płaskim, okrągłym korpusem o złożonej strukturze z małą wnęką środkową. Przysadka mózgowa składa się z trzech części: mózgowej, pośredniej i gruczołowej. Jest to najważniejszy gruczoł wydzielania wewnętrznego, wydzielający różne hormony i regulujący wszystkie inne gruczoły wydzielania wewnętrznego, współczulny i przywspółczulny układ nerwowy.
Mastoid corpus mamillare - leży bezpośrednio za szarym guzkiem i służy jako pośredni odruchowy środek węchowy, który poprzez kompleks formacji łuku łączy się z mózgiem węchowym. Ponadto korpus wyrostka sutkowego jest związany z guzkami wzrokowymi i siatkowatym tworzeniem śródmózgowia. Pies ma sparowane ciało.
Diencephalon
Diencephalon jest układany z niewielkiej liczby komórek w ścianie ogromnej trzeciej komory mózgu, tylko u zwierząt lądowych. A zwłaszcza u ssaków osiąga znaczny rozmiar.
1. Dobrze wyrażona embrionalna płytka opony u wszystkich dorosłych ssaków tworzy nabłonek. Szyszynka jest podstawą trzeciego niesparowanego oka ciemieniowego. Tylko niektóre zwierzęta wodne i gady mają pod skórą pęcherz w kształcie oka. U ssaków szyszynka staje się gruczołem wydzielania wewnętrznego, którego funkcja wciąż nie jest dobrze poznana.
2. Specyficzna i wysoko rozwinięta płodowa płytka dna stanowi podtalaliczną część międzymózgowia. Pochodzi z mózgu epichordalnego, czyli jest połączony z śródmózgiem.
Przysadka mózgowa pochodzi z trzech różnych źródeł. Ektoderma gardła zmienia się w rozgałęziony gruczoł. Światło gruczołu następnie znika, ale pozostają pasma komórek gruczołowych o różnych strukturach, otoczone dużą liczbą naczyń. U zwierząt lądowych część nerwowa przysadki mózgowej z komórek nerwowych i glejowych oraz włókien nerwowych powstaje ze ściany lejka. Pośrednia część przysadki mózgowej z komórek nabłonkowych jest izolowana. Przysadka jako całość wydziela ponad tuzin różnych hormonów, które dostają się do krwioobiegu z części gruczołowej i do płynu mózgowo-rdzeniowego z części nerwowej i pośredniej. Przysadka mózgowa wchodzi w interakcję z centrami autonomicznymi osadzonymi w ścianach trzeciej komory mózgu.
3. Zarodkowa płytka boczna tworzy wzrokowy boraks-wzgórze - i pęcherzyki oka. Struktura wzgórza obejmuje jądra guzków wzrokowych. Służą one:
a) ośrodki pośrednie wszystkich ścieżek, które prowadzą do kory mózgowej i przewodzą różne impulsy - węchowy, wrażliwy na skórę i mięśnie, smak i u zwierząt lądowych, dodatkowo wizualny i statoakustyczny;
b) środkowy środek wszystkich ścieżek, które biegną od kory płaszcza do różnych części mózgu.
To wyjaśnia, dlaczego wizualne guzki zaczynają formować się wraz z pojawieniem się płaszcza węchowego i osiągają maksymalny rozwój u ssaków w związku z tworzeniem się płaszcza wtórnego. Silny rozwój guzków wzrokowych jest również spowodowany ruchem ośrodków wzrokowych od śródmózgowia do wtórnego płaszcza, z powodu połączenia z móżdżkiem. W wyniku wzbogacenia wzgórza o pośrednie centra rośnie pośrednia masa guzków wzrokowych, które po wprowadzeniu do jamy trzeciej komory mózgu zamienia ją w kanał w kształcie pierścienia. W szarej ścianie ściany trzeciej komory mózgowej znajdują się liczne wyższe podkorowe centra autonomiczne.
Śródmózgowo-śródmózgowia
Składa się z: poczwórnej płytki, czapki nóg lub czapki nóg dużego mózgu. Jama śródmózgowia zamieniła się w akwedukt mózgowy aqueductus cerebri - łączy trzecią i czwartą komorę mózgową. Centralny szary rdzeń opony jest osadzony w ścianach wodociągu.
1. płytka kwadrupolowo-tekturowa - reprezentuje grzbietową część śródmózgowia, leży ogonowo od guzków wzrokowych i nosowo od móżdżku. Płytka składa się ze sparowanych nosowo-ogonowych wzgórz-colliculi nasalis et caudalis. Wzgórza są oddzielone poprzecznymi i środkowymi rowkami. Z powierzchni czteroczęściowa płytka jest pokryta białą substancją mózgową, pod którą znajduje się szara substancja mózgowa, na wzgórzach nosowych jest podkorowym centrum różnych ścieżek aferentnych, w tym wizualnych, oraz w ogonowych podkorowych ośrodkach słuchowych i równowagi.
Ogólnie rzecz biorąc, płytka poczwórna jest ośrodkiem koordynującym szereg impulsów: węchowy, wzrokowy, równowagowo-słuchowy, ogólna wrażliwość i impulsy z kory mózgowej. Impulsy silnikowe są przekazywane do czerwonego rdzenia, rdzenia kręgowego, a także do głównych mięśni, móżdżku i mostka.
2 w środku śródmózgowia między płytką poczwórną a nogami dużego mózgu pokrycie stopy lub cap-tegmentum pedunculi . Leżą w nim sparowane jądra szarej rdzenia w płaszczyźnie przednich wzgórz:
a) czerwony rdzeń-jądro rdzeń-motor centrum rdzenia kręgowego;
b) jądro nerwu okoruchowego-jądro n. okulomotorius.
c) przywspółczulne jądra Jakubowicza.
Lokalizacja:
d) jądro bloku nerw-jądro n. trochlealis;
d) część jądra piątej pary nerwów.
Formacja siatkowa przechodzi przez całą nasadkę od rdzenia przedłużonego do produktu pośredniego, tworząc ścieżki od rdzenia kręgowego i móżdżku do kwadrupolu, do guzków wzrokowych, a od nich do rdzenia kręgowego.
3. nogi cerebrum-pedunculi cerebri - pojawiają się na podstawowej powierzchni mózgu w postaci dwóch grubych grzbietów między drogami wzrokowymi a mostkiem mózgu. Są one podzielone przez rowek między stopami. Z nóg wyjdź 3 pary nerwów czaszkowych - nerw okoruchowy. Nogi są utworzone przez odprowadzające ścieki, które łączą korę płaszcza z romboidalnym i rdzeniem kręgowym. Dlatego nogi są bardziej rozwinięte u zwierząt, u których kora mózgowa jest silniejsza. U ludzi są one najbardziej rozwinięte.
Rozwój śródmózgowia
Śródmózgowia u niższych zwierząt, podobnie jak w zarodkach wyższych zwierząt, osiągają bardzo znaczny rozmiar. Szary rdzeń embrionalnej blaszki bocznej środkowego pęcherza mózgowego, rosnący, tworzy łuk śródmózgowia, z którego niższe zwierzęta wytwarzają dicollis lub płaty wzrokowe. Początkowo dicollis było najwyższym ośrodkiem koordynującym, ponieważ kończyły się w nim nie tylko nerwy wzrokowe, ale także ścieżki doprowadzające z prążkowia, romboidu i rdzenia kręgowego. Ale gady mają już niektóre włókna, podczas gdy ssaki mają prawie wszystkie włókna nerw wzrokowy przejdź przez wizualne guzki diencefalonu do kory wtórnego płaszcza. Dlatego wizualne płaty pozostają w tyle pod względem wzrostu, a wtórny płaszcz i, odpowiednio, wizualne guzki rosną coraz bardziej.
U zwierząt lądowych centra równowagowo-słuchowe układane są w łuku śródmózgowia, początkowo w postaci formacji mikroskopowych, a następnie makroskopowych. Tylko u ssaków pojawia się kwadrupol zamiast dychotomii. U zwierząt o dobrym słuchu przeważają wzgórza tylno - słuchowe.
Z embrionalnej płyty głównej w brzusznej ścianie środkowego pęcherza mózgowego tworzy się czapka. W wyniku uformowania łuku i czapki jama śródmózgowia zamienia się w akwedukt mózgowy. Czapka składa się z jąder 3 i 4 par nerwów czaszkowych i specjalnych jąder motorycznych czapki. Te ostatnie obejmują czerwone jądro łączące móżdżek z rdzeniem kręgowym oraz jądro śródmiąższowe łączące się przez zwoje wędzidełka z mózgiem węchowym.
U ssaków, w związku z rozwojem wtórnego płaszcza, czapka jest pęknięta brzusznie przez warstwę białej materii mózgowej ze ścieżek prowadzących od kory mózgowej do rombu i rdzenia kręgowego. Ścieżki te tworzą nogi mózgu, których moc odpowiada stopniowi rozwoju kory mózgowej.
Diamentowy rombencefalon
Podzielony na rdzeń przedłużony i kończynę tylną. Mózg składa się z móżdżku i mostka mózgowego. Pomiędzy móżdżkiem a podłużnym mostem znajduje się czwarta komora mózgowa.
A) Medulla oblongata-medulla oblongata
Kontynuuje ogonowo bez zauważalnej granicy rdzenia kręgowego. Na powierzchni podstawy widoczna jest brzuszna środkowa bruzda -fissura mediana ventralis. Po obu stronach przechodzą boczne rowki - bruzda paramedianus - ogonowo wpływają do środkowego rowka brzusznego. Pomiędzy tymi trzema bruzdami znajdują się dwa wąskie grzbiety - piramidy - piramidy - w nich przechodzą piramidalne wiązki przewodzące od kory mózgowej do rdzenia kręgowego. Ponieważ wiązki przewodzące ciągną się do sznurów bocznych i rdzenia kręgowego, a jednocześnie krzyżują się od prawej do lewej i odwrotnie, powstaje krzyż piramida-decussatio pyramidum. Później z piramidy za mostem dochodzą 4 pary nerwów czaszkowych - nerwu odwodzącego. W pobliżu skrzyżowania piramid i 12 par bocznie od niego odchodzi - nerw gnykowy, ale dwa kolejne nerwy, jeden po drugim, bocznie z nerwu gnykowego: 10 nerw para-błędny, bardziej nerw nosowy-9 para-glosowo-gardłowy.
Szara istota rdzenia przedłużonego jest zgrupowana:
a) w oddzielnych jądrach czuciowych i motorycznych, z których wychodzi 5, 6, 7, 9, 10 i 12 par nerwów czaszkowych;
b) do pośrednich jąder wiązek Gaulle'a i Burdacha;
c) w jądrze znajduje się 8 par i związanych z nimi oliwek nosowych i ogonowych-7.
Wśród jąder znajduje się formacja siatki - formatio reticularis - z przeplecionych włókien nerwowych i komórek nerwowych między nimi, które z rdzenia przedłużonego przechodzą do nasadki śródmózgowia i do diencefalonu. Pełni przede wszystkim funkcję asocjacyjną i koordynacyjną między różnymi rdzeniami rombu i śródmózgowia i jest ośrodkiem oddychania oraz układem sercowo-naczyniowym.
Biała rdzeń rdzenia przedłużonego składa się z dużej liczby wiązek ścieżek biegnących od rdzenia kręgowego do różnych części mózgu i odwrotnie.
B) Móżdżek-móżdżek
Ma prawie kulisty kształt, z dwoma rowkami jest podzielony na środkową część - robaka i dwa boczne płaty. Szara rdzeń tworzy kora mózgowa - kora mózgowa - i pojedyncze jądra leżące w centralnie położonej istocie białej móżdżku. Funkcja - koordynacja ruchów.
Powierzchnia kory robaka-robala jest przecięta poprzecznymi rowkami i szczelinami. Dzięki dwóm głównym pęknięciom robak jest ograniczony do płatów przedniego, środkowego i tylnego - lobus rostralis, caudalis et medialis. Każdy z nich jest połączony z nogami móżdżku - przednim, środkowym i tylnym, składającymi się ze ścieżek. Przednie i tylne końce robaka są zagięte w kierunku brzusznym i do siebie, między nimi pozostaje niewielka szczelina - góra namiotu-fastigium. Biała substancja mózgowa robaka-7 na odcinku strzałkowym przypomina gałąź tui, dlatego nazwano ją drzewem żywej altany. W białej mózgu robaka znajduje się namiotowane jądro, jądro fastigii, które jest podkorowym centrum analizatora równowagi.
Za pomocą rdzenia przedłużonego robak łączy się przez ogonowy mózg-żagiel-rąbek rdzenia żołędziowego, a z kwadrupolem przez nosowy żagiel-żuchwę rdzenia żuchwy nasali.
Robak móżdżkowy - półkula półkul mózgowych - podobny do robaka składa się z licznych płatków, z których jeden jest strzępem kłaczków w postaci niewielkiego wyrostka robaka leżącego za boczną nogą. W białym rdzeniu półkul móżdżku znajdują się jądro zębate jądro zębate, które służą jako centra przenoszenia impulsów motorycznych.
Móżdżek łączy się z rdzeniem przedłużonym. Z mostkiem mózgowym - nogami bocznymi i śródmózgiem - nogami nosowymi.
Ogonowe nogi móżdżku-szypułki móżdżku rdzeń przedłużony - w postaci dwóch grzbietów wystają na grzbietowej powierzchni rdzenia przedłużonego. Ścieżki prowadzą przez nie:
a) z rdzenia kręgowego - wiązki Flexig, Gaulle i Burdach z ich jądrami;
b) z jąder nerwu słuchowego;
c) z oliwek ogonowych;
d) z jąder 5.10 i 12 par nerwów czaszkowych.
Nosowe nogi móżdżkowo-ramiennego conjum ctivum cerebelli- idą pod tylnymi wzgórzami czwórki do nóg móżdżku. Nogi nosa leżą na grzbietowej powierzchni rdzenia przedłużonego. Ścieżki prowadzą przez nie:
a) od rdzenia kręgowego do wiązki ślimakowej Govera;
b) od jąder zębatych półkul móżdżku do jądra czerwonego;
c) w jądrze guzków wzrokowych.
Wewnętrzne ścieżki móżdżku są reprezentowane przez włókna między korą mózgową i jej jądrem oraz włókna asocjacyjne łączące oddzielne żyły w płaszczyźnie strzałkowej.
B) Mózg most-pons cerebri
Leży na przednim końcu rdzenia przedłużonego, na granicy z śródmózgiem, w postaci poprzecznego grzbietu, który na końcach zgina się grzbietowo do móżdżku, tworząc boczne nogi móżdżku-ramienno-pontis. Mostek i nogi składają się ze ścieżek łączących jądra mostka-jądro pontis-z jądrem móżdżku. W rdzeniach mostu zaczynają się ścieżki od końca kory mózgowej i ścieżki do półkul móżdżku. 5 par wychodzi przez boczne sekcje mostu - nerw trójdzielny - Najbardziej masywny nerw ze wszystkich nerwów czaszkowych. Ma dwa korzenie: ruch brzuszno - nosowy i grzbietowo - ogonowy. Na ostatnim znajduje się duży zwoje księżycowe. Ogonowo od mostu, również w kierunku poprzecznym, leży trapezoidalne ciało, korpus trapezoideum, w postaci wąskiego i niskiego grzbietu. Tworzą go ścieżki biegnące z jąder nerwu słuchowego. 8 nerwów para-równowagowo-słuchowych i 7 nerwów para-twarzowych wychodzi przez boczne części trapezoidalnego ciała.
D) Czwarty kwartowy komorowo-komorowy mózg
Umieszczony między móżdżkiem a rdzeniem przedłużonym. Łuk jest obsługiwany przez robaka i żagle mózgu, a rdzeń przedłużony jest dnem.
Dno czwartej komory nazywa się romboidalnym fossa-fossa romboidea. Środkowa i dwa boczne rowki u dołu dołu są sparowaną środkową wyniesieniem-wybitne medialne - na którym przedni kopiec - Colliculi facialis - opiera się o boczną szypułkę móżdżku. W obszarze kopca twarzy znajdują się jądra nerwu odwodzącego i twarzy. Na ogonowym końcu uniesienia środkowego znajduje się pole hipoglikozy w okolicy nerwu gnykowego z jądrem o tej samej nazwie.
Jądra 9 i 10 par nerwów wystają bocznie z nerwu gnykowego. Tworzą szare skrzydło ala cinerea. Obszar tylnego końca szarych skrzydeł jest znany jako tatarak długopis.
Bezpośrednio za bocznymi nogami móżdżku i od nich przyśrodkowo pojawiają się przedsionkowe pola przedsionkowe w postaci niewielkich wzniesień. Zawierają jądro przedsionkowe i ślimakowe 8 par nerwów. Jądra ślimaka leżą bardziej bocznie.
Rozwój diamentów
Rdzeń przedłużony rozwija się przez wzrost bocznych i głównych płytek zarodkowej rurki mózgowej. Z powodu rozszerzenia płytek bocznych o tworzący się móżdżek ośrodki ruchowe rdzenia przedłużonego zostały przesunięte w środkowej części, a czuciowe przesunięto w bok.
To samo wynika z przyśrodkowej lokalizacji rdzenia przedłużonego nerwów ruchowych i wrażliwych na boki, a także cech strukturalnych dna czwartej komory mózgu, na grubości których wrażliwe jądra leżą bocznie od silnika.
Struktura rdzenia przedłużonego ogólnie u zwierząt jest makroskopowo raczej jednolita, co tłumaczy się również jego jednolitą funkcją - ośrodkami aparatu skrzelowego i jego pochodnymi, jelitami, ośrodkami narządów dotyku, odczuwania mięśniowo-stawowych, słuchu i równowagi. Ze względu na fakt, że ścieżki łączące mózg z rdzeniem kręgowym przechodzą przez rdzeń przedłużony, jego szary rdzeń jest podzielony na oddzielne jądra, w przeciwieństwie do zwartej szarej rdzeni rdzenia kręgowego.
W rdzeniu przedłużonym rdzeń motoryczny formacji czapki lub siatki, który najpierw pojawia się u ryb i jest najstarszym stowarzyszeniem i centrum motorycznym, zasługuje na szczególną uwagę. U zwierząt lądowych z powodu motorycznego rdzenia nasadki oliwki nosowe i ogonowe tworzą się jako centra asocjacyjne. Nosowe oliwki pojawiają się tylko u zwierząt lądowych, poczynając od płazów. Służą jako pośrednie centrum na drodze od nerwu ślimakowego do guzków nerwu wzrokowego. Wiązka tych włókien tworzy korpus trapezoidalny. Oliwki ogonowe powstają jeszcze później u ptaków i ssaków. Są one połączone z jądrami sznurków grzbietowych, guzkami wzrokowymi, móżdżkiem i rdzeniem kręgowym. W swoich stosunkach oliwki ogonowe są najściślej związane z funkcją utrzymywania równowagi.
Móżdżek rozwija się w związku z funkcją utrzymywania równowagi ciała i napięcia mięśni. Dlatego jest najbardziej wyraźny u zwierząt, które pływają szybko, biegają, skaczą, latają i słabiej u zwierząt poruszających się wolno. W prymitywnej formie móżdżek przedstawia niesparowaną płytkę i biały lub szary rdzeń. Móżdżek kładzie się na środku embrionalnej płytki romboidalnej pokrywy, a przednie i tylne żagle mózgowe są formowane z przedniej i tylnej części tego ostatniego. Płytka móżdżku, rosnąca od przodu do tyłu, wygina się łukowato w kierunku grzbietowym. Ze względu na pojawienie się podłużnych rowków, środkowa część płytki - korpus móżdżku - i sparowane części boczne - uszy móżdżku, są izolowane. Ciało móżdżku i uszy są móżdżkiem. Ciało móżdżku u zwierząt lądowych jest podzielone poprzecznymi rowkami na przednie, środkowe i tylne płaty, na których pojawiają się dodatkowe poprzeczne rowki. Płat przedni związany jest z mięśniami głowy, a płat środkowy i tylny związany z mięśniami tułowia i kończyn.
U ssaków dominuje średni odsetek. Wzdłużne rowki oddzielają środkową niesparowaną część, robaka, od sparowanych płatów bocznych lub półkul móżdżku. Półkule móżdżku należą do móżdżku. Robak zawiera środki skoordynowanych, synchronicznych ruchów tułowia i kończyn. Półkule móżdżku są najbardziej wyraźne u wyższych ssaków, które mają, w większym lub mniejszym stopniu, zdolność do oddzielania ruchów kończyn. Z kolei poprawa tej zdolności zależy od siły kory mózgowej, która jest najwyższym ośrodkiem aktywności nerwowej, oraz od pojawienia się móżdżku z korą mózgową przez jego boczne nogi i mostek.
Półkule móżdżku i most znajdują się tylko u ssaków, a maksymalny rozwój osiągają u naczelnych. Jednorodną funkcję móżdżku u zwierząt tłumaczy się dość jednolitym struktura histologiczna kora, w której wyróżnia się powierzchniową warstwę molekularną, głęboką ziarnistą i środkową warstwę dużych komórek Purkine.
Uszy prymitywnego móżdżku u zwierząt wodnych są związane z narządami równowagi, to znaczy narządami linii bocznej i mięśniami ogona. Wraz z redukcją tych narządów u zwierząt lądowych zmniejsza się również uszy. U ssaków zachowane są od nich strzępy, łączące się z tylnym płatem robaka.
Ontogeneza robaka zależy również od jego funkcji, która jest szczególnie wyraźnie widoczna w ludzkim mózgu. U noworodka jest on nadal bardzo mały i stanowi zaledwie 1 / 16-1 / 18 masy dużego mózgu. Ale współczynniki te zmieniają się wraz ze wzrostem dziecka i jego ruchliwości; u dorosłego stosunek ten jest już inny - 1 / 8-1 / 9 masy mózgu.
Wykaz używanej literatury
1) A.I. Akayevsky „Anatomy of Pets”
2) I.V. Chrustalewa „Anatomia zwierząt”
3) L.V. Antipova, V.S. Slobodyanik, S. M. Suleymanov „Anatomia i histologia zwierząt gospodarskich”
4) A.F. Rimov „Anatomia zwierząt”
5) A.F. Kuznetsova „Przewodnik po medycynie weterynaryjnej”