Opisz główne rodzaje tkanki mięśniowej. Struktura histologiczna tkanki mięśniowej.
Tkanka mięśniowa nazywana jest tkanką, która ma inną budowę i pochodzenie, ale podobną zdolność do wyraźnych skurczów. Zapewniają ruch w przestrzeni całego organizmu jako całości lub jego części (na przykład mięśni szkieletowych) oraz ruch narządów wewnątrz narządu (na przykład serca, języka, jelit).
Komórki wielu tkanek mają właściwość zmiany kształtu, ale w tkance mięśniowej zdolność ta staje się główną funkcją.
Ogólna charakterystyka i klasyfikacja
Główne cechy morfologiczne elementów tkanki mięśniowej są wydłużone, obecność podłużnie zlokalizowanych miofibryli i miofilamentów - specjalnych organelli, które zapewniają kurczliwość, lokalizacja mitochondriów w pobliżu elementów kurczliwych, obecność wtrąceń glikogenu, lipidów i mioglobiny.
Gładka tkanka mięśniowa.
Ta tkanina jest uformowana z mezenchyme. Komórki mięśni gładkich - wydłużone ze spiczastymi końcami w kształcie wrzeciona, pokryte błoną komórkową. Miocyt to komórka jednojądrzasta. Jądro w kształcie pręta zajmuje centralną pozycję w komórce. Zawiera atomizowaną heterochromatynę i jedno lub dwa wyraźnie widoczne jąderka. Centrum komórkowe (cytocentrum) znajduje się w pobliżu jednej z powierzchni jądra. Pozostałe organelle są skoncentrowane na biegunach jądra w kształcie pręta. Dobrze rozwinięte mitochondria, kompleks Golgiego, gładki EPS; rybosomy, granulowany EPS są lekko wyrażone. Cytoplazma miocytów zawiera granulki glikogenu - rezerwę energetyczną komórki.
Miocyty ściśle przylegają do siebie, tworząc warstwy i grupy, oddzielone luźną nieformowaną tkanką łączną. Miofibryle znajdują się w cytoplazmie; poruszają się wzdłuż obwodu komórki wzdłuż jej osi. Składają się z cienkich nitek i są kurczliwym elementem mięśnia.
Na zewnątrz włókno mięśniowe pokryte jest błoną - sarcolemma, składająca się z wewnętrznej i zewnętrznej warstwy. Warstwa wewnętrzna jest plazmolemem, podobnym do błony innych komórek tkankowych. Zewnętrznie - warstwa tkanki łącznej składa się z membrany piwnicy i sąsiadujących struktur włóknistych. Plazmolemma tworzy system wąskich kanalików, które wnikają do włókna mięśniowego.
Włókna tkanki łącznej znajdujące się poza błoną podstawną włókna mięśniowego tworzą endomysium, które jest bogate w naczynia krwionośne i nerwy. Endomysium łączy się z perimisium - błoną pokrywającą grupę włókien mięśniowych. Krążek kilku wiązek mięśni jest połączony z nabłonkiem - najbardziej zewnętrzną błoną tkanki łącznej, która łączy kilka tych wiązek w mięsień - organ charakteryzujący się specyficzną strukturą i funkcją.
Gładka tkanka mięśniowa znajduje się w ścianach żołądka, jelit, macicy, pęcherz, moczowody, oskrzela, a także w średnich i dużych naczyniach krwionośnych. Aktywność mięśni gładkich jest regulowana przez ANS. Skurcze mięśni nie są zgodne z wolą człowieka, dlatego gładką tkankę mięśniową nazywa się mimowolną muskulaturą.
Prążkowana tkanka mięśniowa.
Tkanka mięśni szkieletowych jest kurczliwą tkanką tułowia, głowy, kończyn, gardła, krtani, górnej połowy przełyku, języka i mięśni żucia.
Powstaje z miotomów somitów mezodermy. Jednostką strukturalną jest włókno prążkowane. Ma cylindryczny korpus, jest pokryty powłoką - sarcolem, a cytoplazma nazywa się - sarkoplazma, w której znajduje się wiele jąder i miofibryli. Każdy miofibryl składa się z dysków o innym składzie chemicznym. Miofibryle to kurczliwy aparat włókien mięśniowych.
Całe mięśnie szkieletowe zbudowane są z tkanki mięśni poprzecznie prążkowanych. Mięśnie są dowolne, ponieważ jego skurcz występuje pod wpływem kory mózgowej.
Tkanka mięśniowa serca.
Ten rodzaj tkanki mięśniowej tworzy środkową błonę serca - mięsień sercowy, z natury redukcji odnosi się do mimowolnej, ponieważ nie jest kontrolowany przez wolę zwierzęcia. Rozwija się z trzewiowego splanchnatoma liścia - blaszki miokardialnej.
Tkanka mięśniowa serca składa się z komórek mięśniowych - kardiomiocytów (miocytów sercowych). Miocyty, łącząc się ze sobą na końcach wzdłuż długiej osi komórek, tworzą strukturę podobną do włókna mięśniowego.
Istnieją dwa rodzaje komórek. : typowe komórki kurczliwe i atypowe miocyty sercowestanowiący układ przewodzenia serca.
Typowe komórki mięśniowe pełnić funkcję skurczową; mają kształt prostokątny, 1-2 jądra znajdują się w środku, miofibryle znajdują się na obrzeżach. Pomiędzy sąsiadującymi miocytami znajdują się dyski insercyjne. Za ich pomocą miocyty gromadzą się we włóknach mięśniowych, oddzielonych cienką włóknistą tkanką łączną. Pomiędzy sąsiadującymi włóknami mięśniowymi łączą się włókna, które zapewniają skurcz mięśnia sercowego jako całości.
Układ przewodzenia serca tworzą włókna mięśniowe składające się z nietypowe komórki mięśniowe. Są większe niż kurczliwe, bogatsze w sarkoplazmy, ale gorsze niż miofibryle, które często się przecinają. Jądra są większe i nie zawsze w środku. Włókna układu przewodzącego są otoczone gęstym splotem włókien nerwowych.
Tkanki mięśniowe (łac. Textus muscularis) nazywane są tkankami, które różnią się budową i pochodzeniem, ale mają podobną zdolność do wyraźnych skurczów. Tkanka mięśniowa składa się z wydłużonych komórek, które pobierają podrażnienia układ nerwowy i odpowiedzieć redukcją. Zapewniają ruch w przestrzeni ciała jako całości, jego ruch narządów w ciele (serce, język, jelita itp.) I składają się z włókien mięśniowych. Komórki wielu tkanek mają właściwość zmiany kształtu, ale w tkankach mięśniowych zdolność ta staje się główną funkcją.
Główne objawy morfologiczne elementów tkanki mięśniowej: wydłużony kształt, obecność podłużnie położonych miofibryli i włókien mięśniowych - specjalne organelle, które zapewniają kurczliwość, lokalizacja mitochondriów w pobliżu elementów kurczliwych, obecność wtrąceń glikogenu, lipidów i mioglobiny.
Specjalne organelle skurczowe, miofilamenty lub miofibryle zapewniają skurcz występujący, gdy dwa główne białka fibrylarne - aktyna i miozyna - wchodzą w interakcję z obowiązkowym udziałem jonów wapnia. Mitochondria dostarczają tym procesom energii. Zapas źródeł energii tworzy glikogen i lipidy. Mioglobina jest białkiem, które zapewnia wiązanie tlenu i tworzenie jego rezerwy w momencie skurczu mięśni, gdy naczynia krwionośne są ściśnięte (gwałtownie spada przepływ tlenu).
Właściwości tkanki mięśniowej:
- Pobudliwość
- Przewodnictwo
- Kurczliwość
- Labilność
Rodzaje tkanki mięśniowej:
1. Gładka tkanka mięśniowa
Gładka tkanka mięśniowa składa się z komórek jednojądrzastych - miocytów w kształcie wrzeciona o długości 20-500 mikronów. Ich cytoplazma w mikroskopie świetlnym wygląda jednolicie, bez poprzecznych prążków. Ta tkanka mięśniowa ma specjalne właściwości: powoli kurczy się i rozluźnia, ma automatyzację, jest mimowolna (to znaczy, jej aktywność nie jest kontrolowana przez wolę osoby). Zawarte w ścianach narządy wewnętrzne: naczynia krwionośne i limfatyczne, drogi moczowe, przewód pokarmowy (skurcze ścian żołądka i jelit).
2. Prążkowana tkanka mięśni szkieletowych
Składa się z miocytów o dużej długości (do kilku centymetrów) i średnicy 50-100 mikronów; komórki te są wielojądrowe, zawierają do 100 lub więcej jąder; w mikroskopie świetlnym cytoplazma wygląda jak naprzemienne ciemne i jasne paski. Właściwości tej tkanki mięśniowej to duża skurcz, relaksacja i przypadkowość (to znaczy, jej aktywność jest kontrolowana przez wolę osoby). Ta tkanka mięśniowa jest częścią mięśni szkieletowych, a także ścian gardła, górnej części przełyku, tworzy język, mięśnie okoruchowe. Włókna o długości od 10 do 12 cm.
3. Prążkowana tkanka mięśnia sercowego
Składa się z 1 lub 2 jądrowych kardiomiocytów mających poprzeczne prążkowanie cytoplazmy (wzdłuż obwodu cytolemmy). Kardiomiocyty są rozgałęzione i tworzą wzajemnie połączone połączenia - dyski insercyjne, w których połączona jest ich cytoplazma. Istnieje również inny kontakt międzykomórkowy - anostamozy (hamowanie cytolemii jednej komórki w cytolemacie innej) Ten typ tkanki mięśniowej tworzy mięsień sercowy serca. Rozwija się z płytki mięśnia sercowego (trzewna warstwa splanchnotoma szyi zarodka) Szczególną właściwością tej tkanki jest automatyzacja - zdolność rytmicznego kurczenia się i relaksacji pod wpływem wzbudzenia, które występuje w samych komórkach (typowe kardiomiocyty). Ta tkanka jest mimowolna (atypowe kardiomiocyty). Istnieje trzeci typ kardiomiocytów - kardiomiocyty wydzielnicze (nie mają włókienek). Syntetyzują hormon troponinę, który obniża ciśnienie krwi i rozszerza ściany naczyń krwionośnych.
Błona mięśniowa serca (mięśnia sercowego) - jest utworzona przez komórki jamy mięśniowej, które są połączone w kompleksy mięśniowe lub włókna mięśnia sercowego za pomocą dysków wprowadzających. Tkanka mięśniowa ma zdolność do regeneracji, jest chroniona przez tkankę łączną, tworząc bliznę.
Przekazanie dobrej pracy do bazy wiedzy jest łatwe. Skorzystaj z poniższego formularza
Będą wam bardzo wdzięczni studenci, doktoranci, młodzi naukowcy, którzy wykorzystują bazę wiedzy w swoich badaniach i pracy.
Wysłano na http://www.allbest.ru/
Tkanka mięśniowa łączy zdolność redukcji.
Cechy strukturalne: aparat kurczliwy, który zajmuje znaczną część w cytoplazmie elementów strukturalnych tkanki mięśniowej i składa się z włókien aktyny i miozyny, które tworzą organelle specjalnego przeznaczenia - miofibryle .
Tkanka mięśniowasą grupą tkanek różnego pochodzenia i struktury, połączonych na podstawie wspólnej cechy - wyraźnej zdolności kurczenia się, dzięki której mogą pełnić swoją główną funkcję - poruszania ciałem lub jego częściami w przestrzeni.
Najważniejsze właściwości tkanki mięśniowej.Elementy strukturalne tkanki mięśniowej (komórki, włókna) mają wydłużony kształt i są w stanie kurczyć się z powodu silnego rozwoju aparatu kurczliwego. Ten ostatni charakteryzuje się bardzo uporządkowanym układem aktynai miofiliny miozyny,tworzenie optymalnych warunków dla ich interakcji. Osiąga się to poprzez połączenie struktur kurczliwych ze specjalnymi elementami cytoszkieletu i plazmatemii (sarcolemma)występować funkcja odniesienia. W części tkanki mięśniowej miofilamenty tworzą organelle o szczególnym znaczeniu - miofibryle.Skurcz mięśni wymaga znacznej ilości energii, dlatego w elementach strukturalnych tkanek mięśniowych występuje duża liczba mitochondriów i wtrąceń troficznych (krople lipidów, granulki glikogenu) zawierających substraty - źródła energii. Ponieważ skurcz mięśni występuje z udziałem jonów wapnia, struktury, które dokonują jego akumulacji i wydalania, są dobrze rozwinięte w komórkach mięśniowych i włóknach - agranularnej siateczce endoplazmatycznej (retikulum sarkoplazmatyczne), jaskinia.
Klasyfikacja tkanki mięśniowejna podstawie znaków ich (a) struktury i funkcji (klasyfikacja morfofunkcjonalna)oraz (b) pochodzenie (klasyfikacja histogenetyczna).
Klasyfikacja morfofunkcjonalna tkanki mięśniowej podkreśla prążkowana (prążkowana) tkanka mięśniowai gładka tkanka mięśniowa.Tkanka mięśni prążkowanych jest utworzona przez elementy strukturalne (komórki, włókna), które mają poprzeczne prążkowanie ze względu na specjalne uporządkowane względne ustawienie w nich włókien mięśniowych aktyny i miozyny. Do prążkowanych tkanek mięśniowych należą szkieletowyi tkanka mięśnia sercowego.Gładka tkanka mięśniowa składa się z komórek, które nie mają poprzecznego prążkowania. Najczęstszym rodzajem tej tkanki jest tkanka mięśni gładkich, która jest częścią ściany różnych narządów (oskrzeli, żołądka, jelit, macicy, jajowodu, moczowodu, pęcherza i naczyń krwionośnych).
Histogenetyczna klasyfikacja tkanki mięśniowej identyfikuje trzy główne typy tkanki mięśniowej: somatyczny(tkanka mięśni szkieletowych) koelomiczny(tkanka mięśniowa serca) i mesenchymal(gładka tkanka mięśniowa narządów wewnętrznych), a także dwa dodatkowe: komórki mioepitelialne(zmodyfikowane nabłonkowe komórki kurczliwe w końcowych odcinkach i małe kanały wydalnicze niektórych gruczołów) i elementy mineralne(kurczliwe komórki pochodzenia nerwowego w tęczówce oka).
Szkieletowa prążkowana (prążkowana) tkanka mięśniowaw swojej masie przewyższa wszelkie inne tkanki ciała i jest najczęstszą tkanką mięśniową ludzkiego ciała. Zapewnia ruch ciała i jego części w przestrzeni oraz utrzymanie postawy (część aparatu ruchowego), tworzy mięśnie okoruchowe, mięśnie ściany jamy ustnej, języka, gardła i krtani. Nieszkieletowa trzewna prążkowana tkanka mięśniowa, która znajduje się w górnej jednej trzeciej przełyku, jest częścią zewnętrznego zwieracza odbytu i cewki moczowej.
mięśniowy miocyt szkieletowy
Szkieletowa prążkowana tkanka mięśniowa rozwija się w okresie embrionalnym od miotomysomitty powodujące aktywną rozszczepialność myoblasty- komórki, które są ułożone w łańcuchy i łączą się ze sobą w obszarze końców z formacją kanaliki mięśniowe (miotubule)zamienia się w włókna mięśniowe.Takie struktury utworzone przez pojedynczą gigantyczną cytoplazmy i liczne jądra są tradycyjnie określane w literaturze rosyjskiej jako symplasty(w tym przypadku - myosymplasty),termin ten jednak nie występuje w przyjętej terminologii międzynarodowej. Niektóre mioblasty nie łączą się z innymi, znajdującymi się na powierzchni włókien i powodującymi powstanie miosatellitocyty- małe komórki, które są kambialnymi elementami tkanki mięśni szkieletowych. Tkanka mięśni szkieletowych uformowana w wiązki prążkowane włókna mięśniowe, które są jednostkami strukturalnymi i funkcjonalnymi.
Włókna mięśniowe tkanka mięśni szkieletowych to cylindryczne formacje o zmiennej długości (od milimetrów do 10-30 cm). Ich średnica również różni się znacznie w zależności od przynależności do określonego mięśnia i rodzaju, stanu funkcjonalnego, stopnia obciążenia funkcjonalnego, stanu odżywienia i innych czynników. W mięśniach włókna mięśniowe tworzą wiązki, w których leżą równolegle i odkształcając się wzajemnie, często przybierają nieregularny, wielopłaszczyznowy kształt, który jest szczególnie wyraźnie widoczny w przekrojach poprzecznych. Pomiędzy włóknami mięśniowymi znajdują się cienkie warstwy luźnych włókien tkanka łącznanoszenie naczyń i nerwów - endomysium.Poprzeczne prążkowanie włókien mięśni szkieletowych z powodu naprzemiennego ciemności dyski anizotropowe (pasma A)i jasne dyski izotropowe (pasmaI). Każdy dysk izotropowy jest przecięty na pół cienką ciemnością linia Z - telefragm. Jądra włókna mięśniowego - stosunkowo lekkie, z 1-2 jąderkami, diploidalne, owalne, spłaszczone - leżą na jego obwodzie pod sarcolemma i znajdują się wzdłuż włókna. Na zewnątrz sarcolemma jest pokryta grubą warstwą membrana piwnicyw które wplecione są włókna siatkowe.
Komórki miosatelitarne (komórki miosatelitarne) - małe spłaszczone komórki znajdujące się w płytkich odciskach sarcolemma włókna mięśniowego i pokryte wspólną błoną podstawową (patrz ryc. 88). Jądro miosatelitarne jest gęste, względnie duże, organelle są małe i nieliczne. Komórki te są aktywowane po uszkodzeniu włókien mięśniowych i zapewniają ich regenerację regeneracyjną. Łącząc się z resztą włókna ze zwiększonym obciążeniem, komórki miosatelitarne są zaangażowane w jego przerost.
Myofibryle tworzą skurczowy aparat włókna mięśniowego, znajdują się w sarkoplazmie na całej długości, zajmując środkową część i są wyraźnie widoczne na poprzecznych odcinkach włókien w postaci małych kropek.
Miofibryle mają własne poprzeczne prążkowanie i są tak uporządkowane we włóknie mięśniowym, że izotropowe i anizotropowe dyski różnych miofibryli pokrywają się, powodując poprzeczne prążkowanie całego włókna. Każdy miofibryl składa się z tysięcy powtarzających się kolejno powiązanych ze sobą struktur - sarcomeres.
Sarcomere (myomer)jest strukturalną i funkcjonalną jednostką miofibryli i reprezentuje część znajdującą się między nimi telopermy (linie Z).Obejmuje dysk anizotropowy i dwie połówki dysków izotropowych - po jednej połowie z każdej strony. Sarcomere utworzone przez uporządkowany system gruby (miozyna)i cienkie (actinomyx) miofilamenty.Z grubymi miofilamentami są związane mesophragmus (linia M)i skoncentrowany w anizotropowym dysku,
i cienkie włókna przyczepione do telogramy (linie Z),tworzą dyski izotropowe i częściowo wnikają w dysk anizotropowy między grubymi włóknami aż do światła pasek Hw centrum dysku anizotropowego.
W mięśniach, podobnie jak w innych tkankach, wyróżnia się dwa rodzaje regeneracji - fizjologiczne i naprawcze. Regeneracja fizjologiczna objawia się przerostem włókien mięśniowych, co wyraża się wzrostem ich grubości i równomiernej długości, wzrostem liczby organelli, głównie miofibryli, a także wzrostem liczby jąder, co ostatecznie objawia się wzrostem zdolności funkcjonalnej włókna mięśniowego. Za pomocą metody radioizotopowej stwierdzono, że wzrost liczby jąder komórkowych we włóknach mięśniowych w warunkach przerostu jest osiągany z powodu podziału komórek miosatelitarnych i późniejszego wejścia komórek potomnych do miosymplastu.
Wzrost liczby miofibryli osiąga się poprzez syntezę białek aktyny i miozyny przez wolne rybosomy, a następnie połączenie tych białek w miofilamenty aktyny i miozyny równolegle z odpowiednimi włóknami sarkomeru. W wyniku tego najpierw dochodzi do pogrubienia miofibryli, a następnie ich rozszczepienia i tworzenia się miofibryli potomnych. Ponadto tworzenie nowych włókien mięśniowych aktyny i miozyny jest możliwe nie równolegle, ale od początku do końca z poprzednimi miofibrylami, osiągając w ten sposób ich wydłużenie. Retikulum sarkoplazmatyczne i kanaliki T w przerośniętym włóknie powstają w wyniku proliferacji poprzednich elementów. W niektórych typach trening mięśni głównie czerwony typ włókien mięśniowych (w styrenach) lub biały rodzaj włókien mięśniowych (w sprinterach). Związany z wiekiem przerost włókien mięśniowych objawia się intensywnie wraz z nadejściem aktywności ruchowej organizmu (1-2 lata), co wynika przede wszystkim ze zwiększonej stymulacji nerwowej. W wieku starczym, a także w warunkach niskiego obciążenia mięśni, dochodzi do zaniku organelli specjalnych i ogólnych, ścieńczenia włókien mięśniowych i zmniejszenia ich zdolności funkcjonalnych.
Regeneracja regeneracyjna rozwija się po uszkodzeniu włókien mięśniowych. Ponadto metoda regeneracji zależy od wielkości wady. Ze znacznymi uszkodzeniami w całym włóknie mięśniowym miozatelity w obszarze uszkodzenia i na obszarach przyległych są hamowane, intensywnie namnażają się, a następnie migrują do obszaru defektu włókna mięśniowego, gdzie wyrównują się, tworząc miotube. Późniejsze różnicowanie probówki prowadzi do uzupełnienia defektu i przywrócenia integralności włókna mięśniowego. W warunkach niewielkiej wady włókna mięśniowego na jego końcach, z powodu regeneracji wewnątrzkomórkowych organelli, powstają nerki mięśniowe, które rosną ku sobie, a następnie łączą się, co prowadzi do zamknięcia wady. Jednak regeneracyjną regenerację i przywrócenie integralności włókien mięśniowych można przeprowadzić w określonych warunkach: po pierwsze, z zachowanym unerwieniem motorycznym włókien mięśniowych, a po drugie, jeśli elementy tkanki łącznej (fibroblasty) nie wchodzą w obszar uszkodzenia. W przeciwnym razie blizna tkanki łącznej rozwija się w miejscu uszkodzenia włókna mięśniowego.
Radziecki naukowiec A.N. Student udowodnił możliwość autotransplantacji tkanki mięśni szkieletowych, a nawet całych mięśni w określonych warunkach:
· Mechaniczne mielenie tkanki mięśniowej przeszczepu w celu odhamowania komórek satelitarnych i ich późniejszej proliferacji;
· Umieszczenie rozdrobnionej tkanki w łóżku powięzi;
· Zwijanie włókna nerwu ruchowego do zmiażdżonego przeszczepu;
· Obecność kurczliwych ruchów antagonistów i synergetyków mięśni.
Anatomicznie noworodki mają wszystkie mięśnie szkieletowe, ale w odniesieniu do masy ciała stanowią tylko 23% (u dorosłych 44%). Ilość włókna mięśniowego w mięśniach jest taka sama jak u osoby dorosłej. Jednak mikrostruktura włókien mięśniowych jest inna: włókna mają mniejszą średnicę, mają więcej jąder. Gdy rośnie, następuje pogrubienie i wydłużenie włókien. Wynika to z pogrubienia miofibryli, popychając jądro na obrzeże. Rozmiar włókien mięśniowych stabilizuje się o 20 lat.
Mięśnie u dzieci są bardziej elastyczne niż u dorosłych. Tj. skracają się szybciej podczas skurczu i wydłużają podczas relaksu. Pobudliwość i labilność mięśni noworodków jest niższa niż u dorosłych, ale wzrasta z wiekiem. U noworodków, nawet we śnie, mięśnie są w stanie napięcia. Rozwój różnych grup mięśni przebiega nierównomiernie. Po 4-5 latach mięśnie przedramienia są bardziej rozwinięte, mięśnie ręki pozostają w tyle w rozwoju. Przyspieszone ocieplenie mięśni dłoni następuje po 6-7 latach. Ponadto prostowniki rozwijają się wolniej niż zginacze. Z wiekiem zmienia się stosunek napięcia mięśniowego. We wczesnym dzieciństwie zwiększone napięcie mięśni dłoni, prostowników bioder itp. stopniowo rozkład tonów zostaje znormalizowany.
Serce jako narząd charakteryzuje się zdolnością do regeneracji poprzez przerost regeneracyjny, w którym przywracana jest masa narządu, ale kształt pozostaje zaburzony. Podobne zjawisko obserwuje się po zawale mięśnia sercowego, kiedy masa serca może się zregenerować jako całość, podczas gdy blizna tkanki łącznej tworzy się w miejscu uszkodzenia, ale przerost narządów, tj. formularz jest zepsuty. Występuje nie tylko wzrost wielkości kardiomiocytów, ale także proliferacja głównie w przedsionkach i uszach serca.
Wcześniej uważano, że różnicowanie kardiomiocytów jest nieodwracalnym procesem związanym z całkowitą utratą zdolności tych komórek do podziału. Jednak na obecnym poziomie wiele danych pokazuje, że zróżnicowane kardiomiocyty są zdolne do syntezy DNA i mitozy. W pracach badawczych P.P. Rumyantsev i jego uczniowie wykazali, że po eksperymentalnym zawale mięśnia sercowego lewej komory serca 60-70% kardiomiocytów przedsionkowych wraca do cyklu komórkowego, liczba komórek poliploidalnych wzrasta, ale to nie kompensuje uszkodzenia mięśnia sercowego.
Stwierdzono, że kardiomiocyty są zdolne do podziału mitotycznego (w tym komórek układu przewodzącego). W mięśniu sercowym znajduje się szczególnie wiele jednojądrzastych komórek poliploidalnych o 16-32-krotnej zawartości DNA, ale kardiomiocyty dwuprzewodowe (13-14%) są głównie oktoploidalne.
W procesie regeneracji tkanki mięśnia sercowego kardiomiocyty uczestniczą w procesie hiperplazji i przerostu, ich ploidalność wzrasta, ale poziom proliferacji komórek tkanki łącznej w obszarze uszkodzenia jest 20-40 razy wyższy. W fibroblastach aktywowana jest synteza kolagenu, w wyniku której dochodzi do naprawy poprzez bliznowanie wady. Biologiczna prezentacja takiej adaptacyjnej reakcji tkanki łącznej tłumaczy się żywotnym znaczeniem narządu sercowego, ponieważ opóźnienie zamknięcia wady może doprowadzić do śmierci.
Uważano, że u noworodków i prawdopodobnie we wczesnym dzieciństwie, gdy nadal są zdolne do dzielenia kardiomiocytów, procesom regeneracyjnym towarzyszy wzrost liczby kardiomiocytów. Ponadto u dorosłych regeneracja fizjologiczna odbywa się w mięśniu sercowym głównie przez regenerację wewnątrzkomórkową, bez zwiększania liczby komórek, tj. w mięśniu sercowym osoby dorosłej nie dochodzi do proliferacji kardiomiocytów. Ale ostatnio uzyskano dowody na to, że w zdrowym ludzkim sercu 14 na milion miocytów znajduje się w stanie mitozy, co powoduje cytotomię, tj. liczba komórek nie jest znacząca, ale rośnie.
Zastosowanie nowoczesnych metod biologii komórki w badaniach klinicznych i eksperymentalnych pozwoliło nam wyjaśnić komórkowe i molekularne mechanizmy uszkodzenia i regeneracji mięśnia sercowego. Szczególnie interesujące są dowody na to, że w regionach perinekrotycznych i w zatkanym funkcjonalnie sercu zachodzi synteza embrionalnych białek i peptydów mięśnia sercowego, a także białek syntetyzowanych podczas cyklu komórkowego. Potwierdza to podobieństwo między mechanizmami regeneracji a normalną ontogenezą.
Okazało się również, że zróżnicowane kardiomiocyty w hodowli są zdolne do aktywnego podziału mitotycznego, co można wytłumaczyć nie całkowitą utratą, ale tłumieniem zdolności kardiomiocytów do powrotu do cyklu komórkowego.
Ważnym zadaniem teoretycznej i praktycznej kardiologii jest opracowanie sposobów stymulowania odbudowy uszkodzonego mięśnia sercowego, tj. indukcja regeneracji mięśnia sercowego i zmniejszenie blizny tkanki łącznej. Jeden z obszarów badań zapewnia możliwość przenoszenia genów regulatorowych, które zamieniają fibroblasty blizny w mioblasty lub transfekcji w kardiomiocyty genów kontrolujących wzrost nowych komórek. Innym kierunkiem jest przeniesienie do obszaru uszkodzenia płodowych komórek szkieletowych i mięśnia sercowego, które mogą być zaangażowane w odbudowę mięśnia sercowego. Przeprowadzane są również eksperymenty z przeszczepem mięśni szkieletowych w sercu, wykazujące tworzenie się w mięśniu sercowym kurczących się tkanek i poprawiające wskaźniki czynnościowe mięśnia sercowego. Leczenie za pomocą czynników wzrostu, które mają zarówno bezpośredni, jak i pośredni wpływ na uszkodzone mięsień sercowy, na przykład poprawę angiogenezy, może być obiecujące.
Gładka tkanka mięśniowa
Trzy grupy mięśni gładkich (lub prążkowanych) rozróżnia się według pochodzenia - mezenchymalny, naskórkowy i nerwowy.
Tkanka mięśniowa pochodzenia mezenchymalnego
Histogeneza. Komórki macierzyste i komórki prekursorowe tkanki mięśni gładkich, już określone, migrują do miejsc nałożenia. Różnicując, syntetyzują składniki macierzy i kolagen błony podstawnej, a także elastynę. W ostatecznych komórkach (miocytach) zdolność syntezy jest zmniejszona, ale nie zanika całkowicie.
Jednostką strukturalną i funkcjonalną gładkiej lub niestriatycznej tkanki mięśniowej jest komórka mięśni gładkich lub gładki miocyt to komórka w kształcie wrzeciona o długości 20-500 μm i szerokości 5-8 μm. Jądro komórki ma kształt pręta, znajduje się w jego środkowej części. Kiedy miocyt się kurczy, jego rdzeń wygina się, a nawet skręca. Organelle o ogólnym znaczeniu, wśród których znajduje się wiele mitochondriów, są skoncentrowane w cytoplazmie w pobliżu biegunów jądra. Aparat Golgiego i ziarnisty retikulum endoplazmatyczne są słabo rozwinięte, co wskazuje na niską aktywność funkcji syntetycznych. Rybosomy są w większości zlokalizowane swobodnie.
Włókna aktynowe w cytoplazmie tworzą trójwymiarową sieć, wydłużoną głównie podłużnie, a raczej ukośnie podłużnie. Końce filamentów są łączone razem i za pomocą plazmolematu za pomocą specjalnych białek sieciujących. Obszary te są wyraźnie widoczne na mikrografach elektronowych jako gęste ciała.
Włókna miozyny są w stanie depolimeryzowanym. Monomery miozyny znajdują się obok włókien aktynowych. Sygnał skurczu zwykle pochodzi z włókien nerwowych. Mediator, który wyróżnia się spośród terminali, zmienia stan plazmy. Tworzy wtargnięcia - kaweolę, w której koncentrują się jony wapnia. Caveolae są splecione w kierunku cytoplazmy w postaci bąbelków (tutaj wapń jest uwalniany z bąbelków). Oznacza to zarówno polimeryzację miozyny, jak i interakcję miozyny z aktyną. Włókna aktyny są przemieszczane względem siebie, gęste plamy łączą się, siła jest przenoszona na błonę plazmatyczną, a cała komórka ulega skróceniu. Kiedy zanikają sygnały z układu nerwowego, jony wapnia są usuwane z jaskiń, miozyna ulega depolimeryzacji i rozkładają się miofibryle. Zatem kompleksy aktyna-miozyna występują w gładkich miocytach tylko w okresie skurczu.
Gładkie miocyty znajdują się bez zauważalnych przestrzeni międzykomórkowych i są oddzielone membraną piwnicy. W niektórych obszarach powstają w nim „okna”, więc plazmolemma sąsiednich miocytów zbiega się. Tutaj powstają ogniwa i między komórkami powstają nie tylko wiązania mechaniczne, ale także metaboliczne. Na „osłonach” błony podstawnej między miocytami znajdują się włókna elastyczne i siatkowe, łączące komórki w pojedynczy kompleks tkanek. Włókna siatkowe wnikają w pęknięcia na końcach miocytów, zostają tam utrwalone i przenoszą siłę skurczu komórek na cały ich związek.
Regeneracja Fizjologiczna regeneracja tkanki mięśni gładkich objawia się w warunkach zwiększonego obciążenia funkcjonalnego. Najwyraźniej widać to w błonie mięśniowej macicy podczas ciąży. Taka regeneracja odbywa się nie tyle na poziomie tkanek, co na poziomie komórkowym: miocyty rosną, procesy cytologiczne ulegają aktywacji w cytoplazmie, rośnie liczba włókien mięśniowych (przerost komórek roboczych). Jednak proliferacja komórek (tj. Rozrost) nie jest wykluczona.
Jako część narządów miocyty są łączone w wiązki, między którymi znajdują się cienkie warstwy tkanki łącznej. Siatkowe i elastyczne włókna otaczające miocyty są wplecione w te warstwy. Naczynia krwionośne i włókna nerwowe przechodzą przez warstwy pośrednie. Końce tych ostatnich nie kończą się bezpośrednio na miocytach, ale między nimi. Dlatego po nadejściu impulsu nerwowego mediator rozprzestrzenia się rozproszony, pobudzając wiele komórek jednocześnie. Gładka tkanka mięśniowa pochodzenia mezenchymalnego jest reprezentowana głównie w ścianach naczyń krwionośnych i wielu cewkowych narządach wewnętrznych, a także tworzy pojedyncze małe mięśnie.
Gładka tkanka mięśniowa w określonych narządach ma nierówne właściwości funkcjonalne. Wynika to z faktu, że na powierzchni narządów znajdują się różne receptory dla określonych substancji biologicznie czynnych. Dlatego w przypadku wielu leków ich reakcja nie jest taka sama.
Gładka tkanka mięśniowa naskórka
Komórki mioepitelialne rozwijają się z zarodka naskórka. Występują w gruczole potowym, sutkowym, ślinowym i łzowym i mają wspólne prekursory z gruczołowymi komórkami wydzielniczymi. Komórki mioepitelialne przylegają bezpośrednio do właściwego nabłonka i mają ze sobą wspólną błonę podstawną. Podczas regeneracji te i inne komórki są przywracane ze wspólnych słabo zróżnicowanych prekursorów. Większość komórek mioepitelialnych ma kształt gwiazdy. Komórki te są często nazywane koszem: ich procesy obejmują sekcje końcowe i małe kanały gruczołów. Rdzeń i organelle o ogólnym znaczeniu znajdują się w ciele komórki, a aparat kurczliwy, zorganizowany, podobnie jak w komórkach tkanki mięśniowej typu mezenchymalnego, znajduje się w procesach.
Gładka tkanka mięśniowa pochodzenia nerwowego
Miocyty tej tkanki rozwijają się z komórek podstawy nerwowej jako części wewnętrznej ściany kielicha wzrokowego. Ciała tych komórek znajdują się w nabłonku tylnej powierzchni tęczówki. Każdy z nich ma proces, który przechodzi w grubość tęczówki i leży równolegle do jej powierzchni. W tym procesie występuje aparat kurczliwy, zorganizowany w taki sam sposób, jak we wszystkich gładkich miocytach. W zależności od kierunku procesów (prostopadłego lub równoległego do krawędzi źrenicy) miocyty tworzą dwa mięśnie - zwężającą się i rozszerzającą źrenicę.
Wniosek
Jak już wspomniano, tkanka mięśniowa to grupa tkanek różnego pochodzenia, połączona zasadą kurczliwości: prążkowana (szkieletowa i sercowa), gładka, a także wyspecjalizowane tkanki kurczliwe - nabłonkowo-mięśniowe i neuroglialne, które są częścią tęczówki oka.
Prążkowana tkanka mięśni szkieletowych powstaje z miotomów, które tworzą elementy segmentowanego mezodermy - somitów.
Gładka tkanka mięśniowa ludzi i kręgowców rozwija się jako część pochodnych mezenchymu, a także tkanki wewnętrznej. Jednak wszystkie tkanki mięśniowe charakteryzują się podobną izolacją w zarodku embrionalnym w postaci komórek w kształcie wrzeciona - komórek tworzących mięśnie lub mioblastów.
Skurcz włókien mięśniowych polega na skracaniu miofibryli w obrębie każdego sarkomeru. Grube (miozyna) i cienkie (aktyna) włókna ciągłe, w stanie zrelaksowanym, połączone tylko sekcjami końcowymi, w momencie skurczu wykonują ruchy ślizgowe w kierunku do siebie. Uwalnianie energii niezbędnej do zmniejszenia energii następuje w wyniku konwersji ATP do ADP pod wpływem miozyny. Aktywność enzymatyczna miozyny objawia się pod warunkiem optymalnej zawartości Ca2 +, które gromadzą się w retikulum sarkoplazmatycznym.
Referencje
1. Histologia. Edytowane przez Yu.I. Afanasyeva, N.A. Yurina. M .: „Medicine”, 1999
2. R. Eckert, D. Rendel, J. Augustine „Animal Physiology” - 1 tom M .: „Mir”, 1981
3. K.P. Ryabov „Histologia z podstawami embriologii” Mińsk: „Wyższa szkoła”, 1990
4. Histologia. Pod redakcją Ulumbekov, prof. Yu.A. Chelysheva. M .: 1998
5. Histologia. Pod redakcją V.G. Eliseeva. M .: „Medicine”, 1983
Wysłany na Allbest.ru
...Podobne dokumenty
Cechy strukturalne tkanki mięśniowej. Badanie mechanizmu skurczu mięśni i aparatu transmisyjnego wzbudzenia. Histogeneza i regeneracja tkanki mięśniowej. Zasady kurczliwych, przewodzących i wydzielniczych kardiomiocytów tkanki mięśnia sercowego.
ściągawka dodana 14.11.2010
Badanie rodzajów i funkcji różnych tkanek ludzkich. Zadania nauki o histologii, która bada strukturę tkanek organizmów żywych. Cechy strukturalne nabłonka, nerwu, tkanki mięśniowej i tkanek środowiska wewnętrznego (łączne, szkieletowe i płynowe).
prezentacja dodana 11/08/2013
Badanie cech strukturalnych tkanek zwierzęcych, funkcjonowania i odmian. Przeprowadzenie badania charakterystycznej cechy struktury tkanki łącznej i nerwowej. Struktura nabłonka płaskiego, sześciennego, rzęskowego i gruczołowego. Rodzaje tkanki mięśniowej.
prezentacja, dodano 02/08/2015
Ogólna charakterystyka i związane z wiekiem cechy tkanki chrzęstnej. Rodzaje chrząstki i tkanki kostnej. Ogólna charakterystyka i związane z wiekiem cechy tkanki kostnej. Cechy struktury tkanki mięśniowej w dzieciństwie i starości. Tkanka mięśni szkieletowych.
prezentacja, dodano 02/07/2016
Klasyfikacja tkanek, rodzaje tkanek nabłonkowych, ich struktura i funkcje. Wspomagająca, troficzna i ochronna funkcja tkanek łącznych. Funkcje tkanek nerwowych i mięśniowych. Pojęcie narządów i układów narządów, ich indywidualność, płeć, różnice wieku.
streszczenie, dodano 09/11/2009
Tkanka nabłonkowa, jej zdolność regeneracyjna. Tkanki łączne zaangażowane w utrzymanie homeostazy środowiska wewnętrznego. Schronienie i komórki limfatyczne. Tkanka mięśni prążkowanych i mięśnia sercowego. Funkcje komórek nerwowych i tkanek organizmów zwierzęcych.
streszczenie, dodano 16 stycznia 2015 r
Badanie rodzajów tkanek zwierzęcych, a także ich funkcji. Cechy strukturalne grup nabłonkowych, łącznych, mięśniowych i nerwowych. Określenie położenia każdej grupy i znaczenia dla życia zwierzęcia.
prezentacja, dodano 10/18/2013
Ogólna koncepcja i rodzaje wibracji. Charakterystyka procesów rozciągania (ściskania), ścinania, zginania, skręcania. Mechaniczne właściwości kości i tkanki naczyniowej. Specyfika tkanki mięśniowej, główne tryby pracy mięśni - izometryczny i izotoniczny.
prace kontrolne, dodano 19.03.2014
Komórka jako główna jednostka strukturalna ciała. Opis jego budowy, właściwości życiowych i chemicznych. Struktura i funkcje nabłonka i tkanki łącznej, mięśni i tkanki nerwowej. Narządy i lista układów narządów ludzkich, ich cel i funkcje.
prezentacja, dodano 19.04.2012
Fizjologia i biochemia aktywności mięśni jako ważny składnik metabolizmu w organizmie. Rodzaje tkanek mięśniowych i odpowiednio mięśnie, które różnią się budową włókien mięśniowych, naturą unerwienia. Efekt aktywności fizycznej o różnym natężeniu.
Rozdział 9. TKANKI MIĘŚNIOWE
Rozdział 9. TKANKI MIĘŚNIOWE
Tkanka mięśniowa (textus muscularis)nazywane tkaninami, które różnią się budową i pochodzeniem, ale mają podobną zdolność do redukcji. Zapewniają ruch w przestrzeni ciała jako całości, jego części i ruch narządów w ciele (serce, język, jelita itp.).
Komórki wielu tkanek mają właściwość kurczenia się ze zmianą kształtu, ale w tkance mięśniowej zdolność ta staje się główną funkcją.
9.1 OGÓLNA CHARAKTERYSTYKA MORFOFUNKCYJNA I KLASYFIKACJA
Główne cechy morfologiczne elementów tkanki mięśniowej są wydłużone, obecność podłużnie zlokalizowanych miofibryli i miofilamentów - specjalnych organelli, które zapewniają kurczliwość, lokalizacja mitochondriów w pobliżu elementów kurczliwych, obecność wtrąceń glikogenu, lipidów i mioglobiny.
Specjalne organelle skurczowe - miofilamenty lub miofibryle, zapewniają skurcz występujący, gdy dwa główne białka fibrylarne - aktyna i miozyna - wchodzą w interakcje z nimi, przy obowiązkowym udziale jonów wapnia. Mitochondria dostarczają tym procesom energii. Zapas źródeł energii tworzy glikogen i lipidy. Mioglobina jest białkiem, które zapewnia wiązanie tlenu i tworzenie jego rezerwy w momencie skurczu mięśni, gdy naczynia krwionośne są ściśnięte (przepływ tlenu dramatycznie spada).
KlasyfikacjaKlasyfikacja tkanki mięśniowej opiera się na dwóch zasadach - morfofunkcjonalnym i histogenetycznym. Zgodnie z zasadą morfofunkcjonalną, w zależności od struktury organelli skurczu, tkanka mięśniowa dzieli się na dwie podgrupy.
Pierwsza podgrupa- prążkowana (prążkowana) tkanka mięśniowa (textus muscularis striatus).W cytoplazmie ich pierwiastków filozy miozyny
jesteś stale spolimeryzowany, tworzysz istniejące miofibryle z filamentami aktynowymi Te ostatnie są zorganizowane w charakterystyczne kompleksy - sarcomeres.W sąsiednich miofibrylach strukturalne podjednostki sarcomeres znajdują się na tym samym poziomie i tworzą prążkowanie krzyżowe.
Druga podgrupa- gładka (nie prążkowana) tkanka mięśniowa (textus muscularis nonstriatus).Te tkanki charakteryzują się tym, że bez skurczu włókna miozyny ulegają depolimeryzacji. W obecności jonów wapnia polimeryzują i oddziałują z włóknami aktynowymi. Utworzone w tym przypadku miofibryle nie mają poprzecznego prążkowania: w specjalnych kolorach są reprezentowane przez (gładkie) nici jednolicie barwione na całej długości.
Zgodnie z zasadą histogenetyczną, w zależności od źródeł rozwoju (embrionalne primordia), tkanka mięśniowa i elementy mięśniowe dzielą się na: somatyczną (miotom), koelomiczną (z płytki mioepikardialnej arkusza trzewnego splanchnotomu), mezenchymalną (z zarodka desmen w mezenchymie (nerwowy), nerwowy rura nerwowa), naskórkowy (z ektodermy skórnej i płytki przedskórnej).
9.2 PASKI MIĘŚNIOWE TKANKI
Istnieją dwie główne odmiany tkanek prążkowanych (prążkowanych) - szkieletowa (miotom) i sercowa (koelomiczna).
9.2.1 Tkanka mięśni szkieletowych
Histogeneza.Źródło rozwoju elementów szkieletowej (somatycznej) tkanki prążkowanej (textus muscularis striatus sceletalis)to komórki macierzyste miotomów - promioblasty. Niektóre z nich różnicują się na miejscu i uczestniczą w tworzeniu tzw. Mięśni autochtonicznych. Inne komórki migrują z miotomów do mezenchymu. Są już określone, chociaż na zewnątrz nie różnią się od innych komórek mezenchymu. Ich zróżnicowanie trwa w miejscach, w których układane są inne mięśnie ciała. Podczas różnicowania powstają dwie linie komórkowe. Komórki jednej z linii łączą się, tworząc wydłużone symplasty - rurki mięśniowe (miotuby). W nich różnicowanie specjalnych organelli - miofibryli (ryc. 9.1). W tym czasie w miotubach zanotowano dobrze rozwinięty ziarnisty retikulum endoplazmatyczne. Miofibryle znajdują się najpierw pod plazmolemem, a następnie wypełniają większość miotubów. Przeciwnie, jądra centralne są przenoszone na peryferia. Centra komórkowe i mikrotubule całkowicie znikają. Granulowane endo
Ryc. 9.1Histogeneza szkieletowej tkanki mięśniowej (według A. A. Klishova):
ale- promioblasty; b- myosymplast; w- rurka mięśniowa; g- dojrzały mięsień
włókno. 1 - miosatelita; 2 - rdzeń miosymplastyczny; 3 - miofibryle
sieć plazmowa jest znacznie zmniejszona. Takie ostateczne struktury nazywa się miosymplastami.
Komórki drugiej linii pozostają niezależne i różnicują się w komórki miosatelitarne. Komórki te znajdują się na powierzchni miosymplastów. Komórki miosatelitarne, rozmnażając się, łączą się z miosymplastami, uczestnicząc w ten sposób w tworzeniu optymalnego jądra
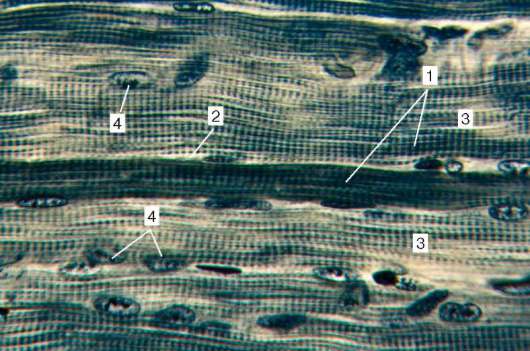
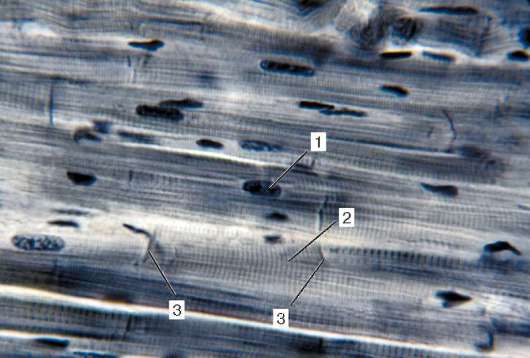
Ryc. 9.2Struktura prążkowanej tkanki mięśniowej (mikrografia):
1 - włókna mięśniowe; 2 - sarcolemma; 3 - sarkoplazma i miofibryle; 4 - rdzenie
myosymplast. Barwienie - żelazo hematoksylina
związek sarkoplazmatyczny niezbędny do syntezy specyficznych białek symplastycznych.
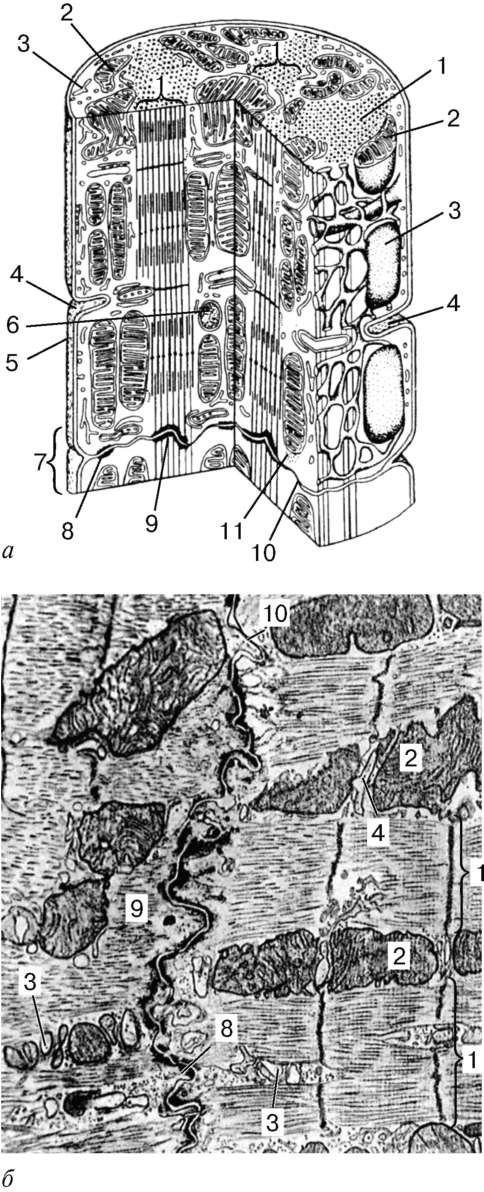
StrukturaGłówną jednostką strukturalną szkieletowej tkanki mięśniowej jest włókno mięśniowe, składające się z miosymplastów i miosatelocytów, pokryte wspólną błoną podstawową (ryc. 9.2-9.4). Długość całego włókna można zmierzyć w centymetrach o grubości 50-100 mikronów. Kompleks składający się z plazmolemii miosymplastu i błony podstawnej nazywa się sarcolemma.
Struktura miosymplastu.Myosymplast ma wiele podłużnych jąder znajdujących się bezpośrednio pod plazmolemem. Ich liczba w jednym symplastie może osiągnąć kilkadziesiąt tysięcy (patrz ryc. 9.2). Na biegunach jąder znajdują się organelle o ogólnym znaczeniu - kompleks Golgiego i małe fragmenty agranularnej retikulum endoplazmatycznego. Miofibryle wypełniają główną część miosymplastu i znajdują się wzdłużnie (patrz ryc. 9.3).
Sarcomere- jednostka strukturalna miofibryli. Każdy miofibryl ma poprzeczne ciemne i jasne dyski o nierównym załamaniu (anizotropowe dyski A i izotropowe dyski I). Każdy miofibryl jest otoczony pętlami agranularnego retikulum endoplazmatycznego - retikulum sarkoplazmatycznego - usytuowanego wzdłużnie i zespolonego między sobą. Sąsiadujące sarcomeres mają wspólną strukturę graniczną - linię Z (ryc. 9.5). Jest zbudowany w postaci sieci białkowych cząsteczek fibrylarnych, wśród których alfa-aktynina odgrywa znaczącą rolę. Końce włókien aktynowych są podłączone do tej sieci. Z sąsiednich linii Z włókna aktynowe są kierowane do środka sarkomeru, ale nie docierają do jego środka. Włókna aktynowe połączone z włóknami Z-line i miozynowymi
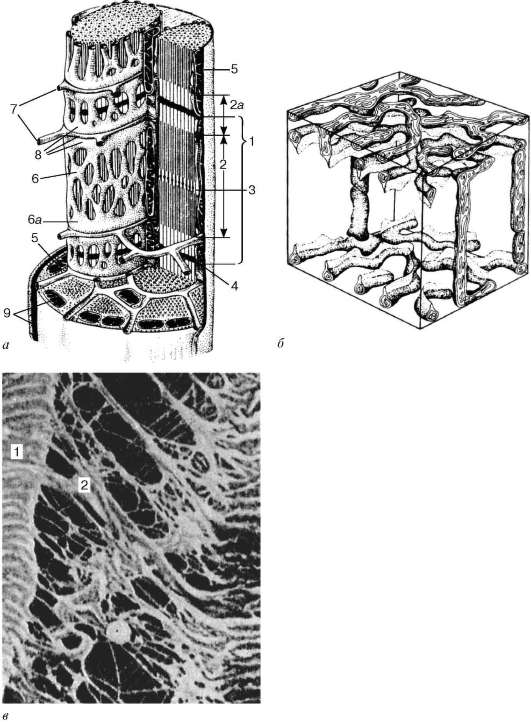
Ryc. 9.3Schemat struktury ultramikroskopowej miosymplastu (zgodnie z R.V. Krstic, ze zmianami) (a): 1 - sarkomer; 2 - dysk anizotropowy (linia A); 2a - dysk izotropowy (ścieżka I); 3 - linia M (mezofragma) na środku dysku anizotropowego; 4 - linia Z (telofragm) na środku dysku izotropowego; 5 - mitochondria; 6 - retikulum sarkoplazmiczne; 6a - zbiornik końcowy; 7 - rurka poprzeczna (rurka T); 8 - triada; 9 - sarcolemma; b- Układ przestrzenny mitochondriów w symplastie. Górne i dolne płaszczyzny figury są ograniczone anizotropowym dyskiem sarko-miary (według L. E. Bakeeva, V. P. Skulachev, Yu. S. Chentsov); w- endomysium. Skaningowy mikrograf elektronowy, powiększenie 2600 (opracowanie Yu. A. Choroszkow): 1 - włókna mięśniowe; 2 - fibryle kolagenowe
fibrylarne nierozciągliwe cząsteczki nebuliny. W środku ciemnego dysku sarkomeru znajduje się sieć zbudowana z miomyozyny. W przekroju tworzy linię M. W węzłach tej linii M końce włókien miozyny są nieruchome. Ich pozostałe końce są skierowane w stronę linii Z i
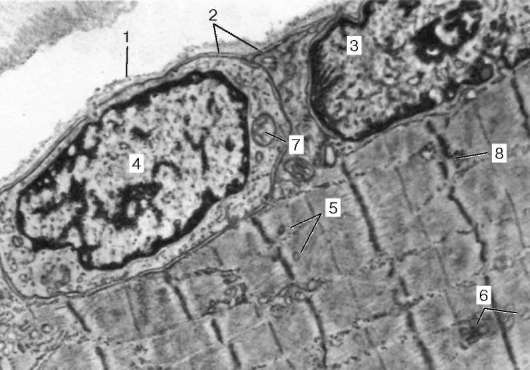
Ryc. 9.4Pole powierzchni miosymplastu i miosatelity. Mikrografia elektronowa, powiększenie 10000 (opracowanie V. L. Goryachkina i S. L. Kuznetsova): 1 - błona podstawowa; 2 - plazmolemma; 3 - rdzeń miosymplastyczny; 4 - jądro miok Tellitocytu; 5 - miofibryle; 6 - kanaliki agranularnej sieci endoplazmatycznej (sarkoplazmatycznej); 7 - mitochondria; 8 - glikogen
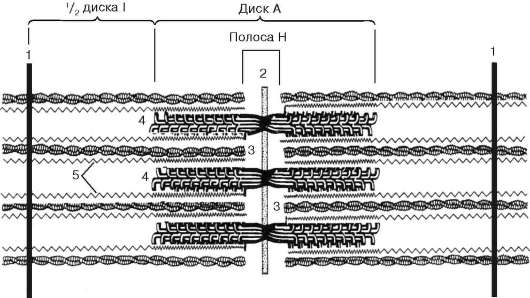
Ryc. 9.5Sarcomere (schemat):
1 - linia Z; 2 - linia M; 3 - włókna aktynowe; 4 - filamenty miozyny; 5 - cząsteczki fibrylarnej tyny (zgodnie z B. Alberts, D. Bray, J. Lewis i in., Z późniejszymi zmianami)
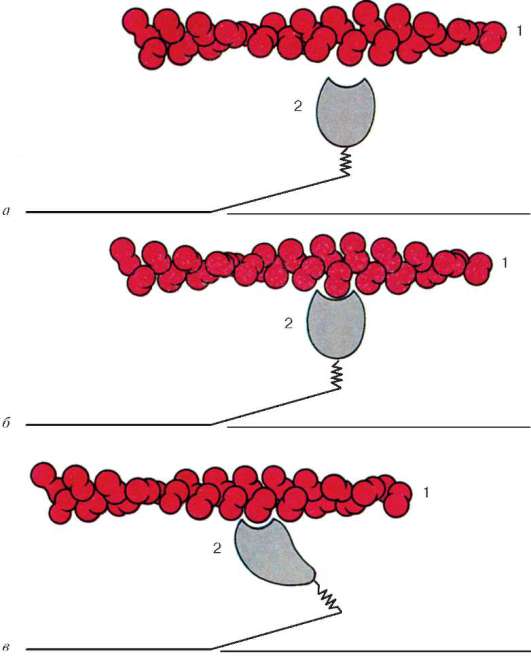
Ryc. 9,6Zmiany konformacyjne pociągające za sobą wzajemne wypieranie włókien aktyny i miozyny:
a-b- sekwencyjne zmiany w relacjach przestrzennych. 1 - aktyna; 2 - głowa cząsteczki miozyny (zgodnie z B. Alberts, D. Bray, J. Lewis i in., Z późniejszymi zmianami)
znajdują się między włóknami aktynowymi, ale również nie osiągają samych linii Z. Jednocześnie końce te są ustalone w odniesieniu do linii Z przez gigantyczne cząsteczki białka tyniny o dużej wytrzymałości na rozciąganie.
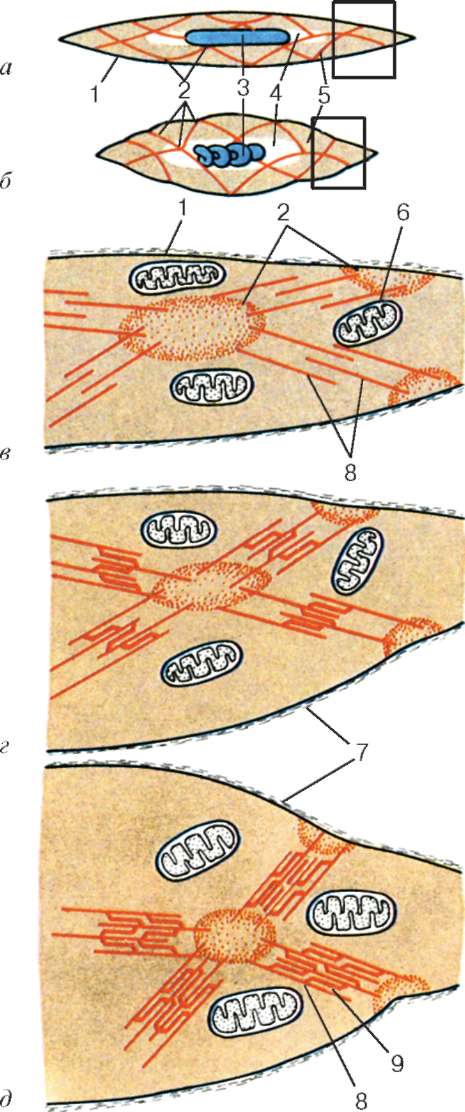
Cząsteczki miozyny mają długi ogon i dwie głowy na jednym końcu. Wraz ze wzrostem stężenia jonów wapnia w obszarze mocowania głów (sekcja zawiasowa) cząsteczka zmienia swoją konfigurację (ryc. 9.6). W tym samym czasie (ponieważ między filamentami miozyny występują aktyna), głowy miozyny wiążą się z aktyną (z udziałem białek pomocniczych - tropomyozyny i troponiny). Następnie głowa miozyny przechyla się i ciągnie cząsteczkę aktyny wzdłuż linii M. Linie Z zbiegają się, mięsak jest skrócony.
Sieci alfa-aktyninowe linii Z sąsiednich miofibryli są połączone ze sobą włóknami pośrednimi. Zbliżają się do wewnętrznej powierzchni plazmy i są unieruchomione w jej warstwie korowej, tak że sarcomeres wszystkich miofibryli znajdują się na tym samym poziomie. Obserwowane pod mikroskopem powoduje wrażenie poprzecznego prążkowania całego włókna.
Źródłem jonów wapnia są zbiorniki agranularnej retikulum endoplazmatycznego. Wydłużają się one wzdłuż miofibryli w pobliżu każdego sarkomeru i tworzą siateczkę sarkoplazmatyczną. To w nim jony wapnia gromadzą się, gdy miosymplast jest w stanie zrelaksowanym. Na poziomie linii Z (u płazów) lub na granicy dysków A i I (u ssaków) kanaliki sieci zmieniają kierunek i są poprzeczne, tworząc rozszerzone czołowe lub boczne (L) zbiorniki.
Z powierzchni w głąb miosymplastu plazmolemma tworzy długie kanaliki rozciągające się poprzecznie (kanaliki T) na poziomie granic między ciemnymi i jasnymi dyskami. Kiedy miosymplast odbiera sygnał o początku skurczu, porusza się wzdłuż plazmolemmy jako potencjał czynnościowy i rozprzestrzenia się na błonę T-rurki. Ponieważ błona ta znajduje się blisko membran siateczki sarkoplazmatycznej, jej stan się zmienia, wapń jest uwalniany ze zbiorników sieci i wchodzi w interakcję z kompleksami aktyna-miozyna (kurczą się). Kiedy potencjał czynnościowy zanika, wapń ponownie gromadzi się w kanalikach sieci, a skurcz miofibryli ustaje. Energia jest potrzebna do opracowania wysiłku redukcyjnego. Uwalnia się go poprzez konwersję ATP na ADP. Rolę ATPazy pełni miozyna. Źródłem ATP są głównie mitochondria, dlatego znajdują się one bezpośrednio między miofibrylami.
Ważną rolę w aktywności miosymplastów odgrywają inkluzje mioglobiny i glikogenu. Glikogen służy jako źródło energii, niezbędnej nie tylko do wykonywania pracy mięśniowej, ale także do utrzymania równowagi cieplnej całego organizmu. Mioglobina wiąże tlen, gdy mięsień jest rozluźniony, a krew swobodnie przepływa przez małe naczynia krwionośne. Podczas skurczu mięśni naczynia krwionośne są ściskane, a zgromadzony tlen jest uwalniany i bierze udział w reakcjach biochemicznych.
Komórki miosatelitarne.Te słabo zróżnicowane komórki są źródłem regeneracji tkanki mięśniowej. Są one przylegające do powierzchni miosymplastu, tak że ich plazmolemma styka się (patrz ryc. 9.1, 9.4). Myosatellitocyty są jednojądrowe, ich ciemne jądra są owalne i mniejsze niż w symplastach. Posiadają wszystkie organelle o ogólnym znaczeniu (w tym centrum komórkowe).
Rodzaje włókien mięśniowych.Różne mięśnie (takie jak narządy) funkcjonują w różnych warunkach biomechanicznych. Dlatego włókna mięśniowe w składzie różnych mięśni mają różną siłę, szybkość i czas trwania skurczu, a także zmęczenie. Aktywność enzymów w nich jest różna i występują one w różnych formach izomerycznych. Zawartość enzymów oddechowych - glikolitycznych i utleniających - jest również w nich różna.
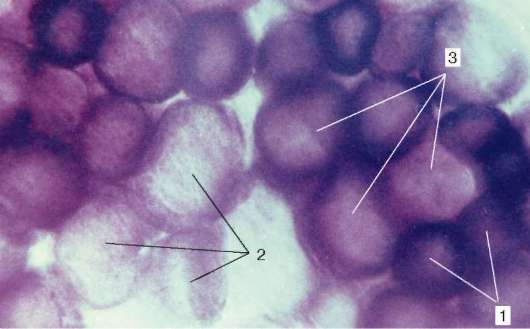
Ryc. 9.7Aktywność dehydrogenazy bursztynianowej we włóknach różnych typów (lek V. F. Chetvergova, przetwarzanie według Nakhlasa i in.): 1 - wysoka; 2 - niski; 3 - średni
Zgodnie ze stosunkiem miofibryli, mitochondriów i mioglobiny rozróżnia się białe, czerwone i pośrednie włókna. Ze względu na cechy funkcjonalne włókna mięśniowe dzielą się na szybkie, wolne i pośrednie, co zależy od organizacji molekularnej miozyny. Wśród jego izoform są dwa główne - „szybki” i „wolny”. Podczas oceny reakcji histochemicznych są one identyfikowane przez aktywność ATPazy. Aktywność enzymów oddechowych również koreluje z tymi właściwościami. Zazwyczaj procesy glikolityczne przeważają w szybkich włóknach, są bogate w glikogen, mają mniej mioglobiny, dlatego nazywane są białymi. Przeciwnie, w wolnych włóknach aktywność enzymów utleniających jest wyższa; są one bogatsze w mioglobinę i wyglądają bardziej na czerwono.
Wraz z białymi i czerwonymi istnieją włókna pośrednie. W większości mięśni szkieletowych włóknami różnych typów histochemicznych są mozaiki (ryc. 9.7).
Właściwości włókien mięśniowych zmieniają się wraz ze zmieniającymi się obciążeniami - sportowymi, zawodowymi, a także w ekstremalnych warunkach (nieważkości). Po powrocie do normalnych czynności takie zmiany są odwracalne. W niektórych chorobach (zanik mięśni, dystrofia, konsekwencje odnerwienia) włókna mięśniowe o różnych właściwościach początkowych zmieniają się różnie. Pozwala to wyjaśnić diagnozę, dla której badają próbki biopsji mięśni szkieletowych.
RegeneracjaJądra miosymplastów nie mogą się dzielić, ponieważ w sarkoplazmie nie ma centrów komórkowych. Elementy kambialne służą komórki miosatelitarne.Podczas gdy ciało rośnie, dzielą się, a komórki potomne łączą się z miosymplastami. Pod koniec wzrostu rozmnażanie komórek miosatelitarnych zanika. Po uszkodzeniu włókna mięśniowego do pewnego stopnia z miejsca urazu zostaje ono zniszczone i jego fragment
jesteś fagocytozowany przez makrofagi. Przywrócenie tkanki odbywa się za pomocą dwóch mechanizmów: kompensacyjnego przerostu samego symplastu i proliferacji komórek miosatelitarnych. W symplastie aktywowany jest ziarnisty retikulum endoplazmatyczny i kompleks Golgiego. Następuje synteza substancji niezbędnych do przywrócenia sarkoplazmy i miofibryli, a także montaż membran, dzięki czemu przywrócona zostaje integralność lematu plazmy. W tym przypadku uszkodzony koniec miosymplastu pogrubia się, tworząc nerkę mięśniową. Komórki miosatelitarne, które przeżywają w pobliżu zmiany są podzielone. Niektóre z nich migrują do nerki mięśniowej i integrują się z nimi, podczas gdy inne łączą się (podobnie jak mioblasty podczas histogenezy) i tworzą nowe miotuby, które przekształcają się we włókna mięśniowe.
9.2.2 Mięśnie szkieletowe jako narząd
Przenoszenie wysiłków skurczowych na szkielet odbywa się za pomocą ścięgien lub poprzez przyczepienie mięśni bezpośrednio do okostnej. Na końcu każdego włókna mięśniowego plazmolemma tworzy głębokie wąskie wybrzuszenia. Cienkie włókna kolagenowe wnikają w nie ze ścięgna lub okostnej. Te ostatnie są spiralnie plecione włóknami siatkowymi. Końce włókien idą do membrany piwnicy, wchodzą do niej, zawracają, a przy wyjściu ponownie splatają włókna kolagenowe tkanki łącznej.
Pomiędzy włóknami mięśniowymi są cienkie warstwy luźnej włóknistej tkanki łącznej - endomysium.Wplecione są w nią włókna kolagenowe zewnętrznego liścia błony podstawnej (patrz ryc. 9.3, c), co pomaga łączyć wysiłki przy jednoczesnym zmniejszeniu miosymplastów. Tworzą się grubsze warstwy luźnej tkanki łącznej otaczającej kilka włókien mięśniowych perimisiumi dzieląc mięsień na wiązki. Kilka wiązek łączy się w większe grupy, oddzielone grubszymi warstwami tkanki łącznej. Nazywa się tkankę łączną otaczającą powierzchnię mięśnia epimysium.
Unaczynienie.Tętnice wchodzą do mięśnia i rozprzestrzeniają się przez warstwy tkanki łącznej, stopniowo przerzedzając. Gałęzie piątego i szóstego rzędu tworzą tętniczki w okostnej. Kapilary znajdują się w endomysium. Idą wzdłuż włókien mięśniowych, zespalając się ze sobą. Żylniki, żyły i naczynia limfatyczne przechodzą w pobliżu naczyń dostawczych. Jak zwykle istnieje wiele komórek tucznych w pobliżu naczyń zaangażowanych w regulację przepuszczalności ściany naczyń.
Innervation.Mięsień ujawnił mielinowany eferentny (motoryczny), aferentny (wrażliwy), a także niemielinowane wegetatywne włókna nerwowe. Scion komórka nerwowaprzynosząc impuls z neuronu ruchowego rdzeń kręgowygałęzie w perimisia. Każdy z jego odgałęzień przenika przez membranę piwnicy i tworzy końcówki na powierzchni plazmolemmy na powierzchni symplastu, uczestnicząc w organizacji tzw. Płytki motorycznej (patrz rozdział 10, ryc. 10.18). Przy przyjęciu
Ryc. 9,8Fragment wrzeciona mięśniowego zawierający włókna mięśniowe z łańcuchem jądrowym (a) i workiem jądrowym (b) (schemat według G. S. Katinas): 1 - jądro; 2 - miofibryle (organelli o znaczeniu ogólnym nie pokazano)
acetylocholina jest uwalniana z końców impulsu nerwowego - mediatora, który powoduje wzbudzenie (potencjał czynnościowy), które rozprzestrzenia się stąd poprzez plazmatolemię miosymplastyczną.
Zatem każde włókno mięśniowe jest unerwione niezależnie i jest otoczone siecią naczyń krwionośnych, tworząc kompleks zwany myon.
Nazywa się grupę włókien mięśniowych unerwionych przez jeden neuron ruchowy jednostka nerwowo-mięśniowa.Włókna mięśniowe należące do jednej jednostki nerwowo-mięśniowej nie leżą w pobliżu, ale znajdują się mozaiki wśród włókien należących do innych jednostek.
Wrażliwe zakończenia nerwowe nie są umiejscowione na pracujących włóknach mięśniowych, ale są powiązane ze specjalistycznymi włóknami mięśniowymi w tak zwanych włóknach mięśniowych.
tenakh (z wklęsłymi włóknami mięśniowymi), które znajdują się w okolicy okostnej.
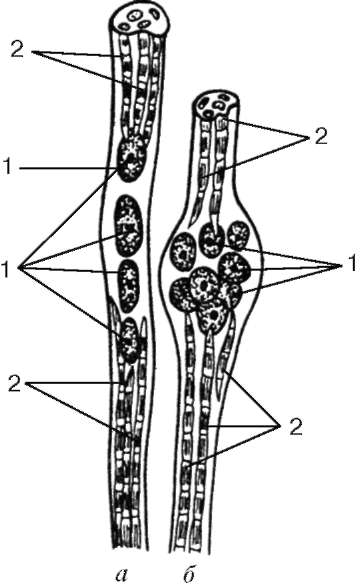
Włókna mięśnia śródgałkowego.Włókna mięśnia śródgałkowego wrzecion są znacznie cieńsze niż pracownicy. Istnieją dwa typy: włókno z workiem jądrowym i włókno z łańcuchem jądrowym (ryc. 9.8). Jądra w tych i innych są zaokrąglone i znajdują się w grubości symplastu, a nie na jego powierzchni. We włóknach z workiem jądrowym jądra symplastu tworzą skupiska w pogrubionej środkowej części. We włóknach z łańcuchem jądrowym w środkowej części symplastu nie powstaje pogrubienie, jądra leżą tutaj wzdłużnie jeden za drugim. Organelle o ogólnym znaczeniu znajdują się w pobliżu skupisk jąder.
Miofibryle znajdują się na końcach symplastów. Włókno Sarcolemma łączy się z kapsułką wrzeciona nerwowo-mięśniowego, składającą się z gęstej włóknistej tkanki łącznej. Każde włókno mięśniowe wrzeciona jest spiralnie splecione z końcem włókna nerwu czuciowego. W wyniku zmniejszenia lub rozluźnienia pracujących włókien mięśniowych zmienia się napięcie torebki tkanki łącznej wrzeciona, a zatem zmienia się ton włókien mięśniowych śródręcza. W wyniku tego wrażliwe zakończenia nerwowe są wzbudzane wokół nich, a impulsowe nerwy pojawiają się w obszarze końcowym. Każdy myosymplast ma również własną tablicę motoryczną. Właśnie dlatego włókna mięśnia śródręcznego są stale napięte, dostosowując się do długości brzucha mięśni jako całości.
9.2.3 Tkanka mięśniowa serca
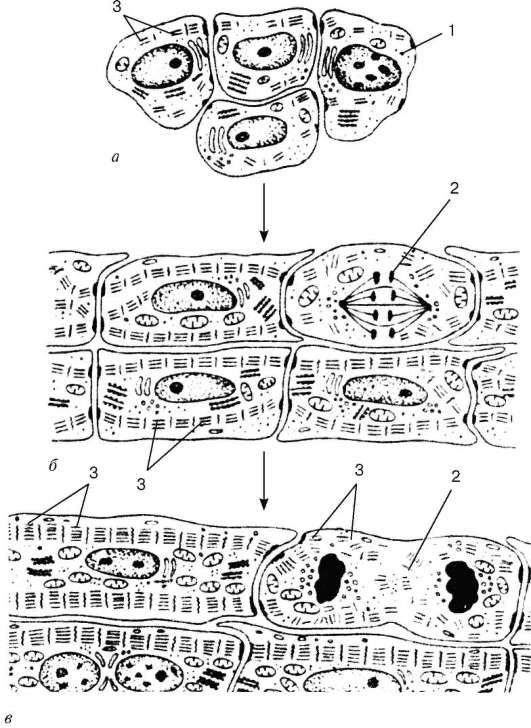
Histogeneza i typy komórek.Źródła rozwoju mięśnia prążkowanego serca (textus muscularis striatus cardiacus)- symetryczne odcinki trzewnego liścia splanchnotome w odcinku szyjnym zarodka - płytki miokardialne.Spośród nich różnicują się także komórki nabłonka międzybłoniaka. Oryginalne komórki tkanki mięśnia sercowego - wybuchy cardiomyo- charakteryzuje się szeregiem znaków: komórki są spłaszczone, zawierają duże jądro, lekką cytoplazmy, ubogie w rybosomy i mitochondria. W przyszłości rozwija się kompleks Golgi, granulowana sieć endo-plazmowa. Struktury włókienkowe znajdują się w kardiomioblastach, ale nie ma miofibryli. Komórki mają wysoki potencjał proliferacyjny.
Po serii cykli mitotycznych kardiomioblasty różnicują się kardiomiocytyw której rozpoczyna się sarkomerogeneza (ryc. 9.9). W cytoplazmie kardiomiocytów wzrasta liczba polisomów i kanalików ziarnistej retikulum endoplazmatycznego, gromadzą się granulki glikogenu i zwiększa się objętość kompleksu aktomyozyny. Kardiomiocyty kurczą się, ale nie tracą zdolności do dalszego namnażania i różnicowania. Rozwój aparatu kurczliwego w późnym okresie embrionalnym i pourodzeniowym następuje przez dodanie nowych sarcomeres i nakładanie warstw nowo zsyntetyzowanych włókien mięśniowych.
Zróżnicowaniu kardiomiocytów towarzyszy wzrost liczby mitochondriów, ich rozmieszczenie na biegunach jąder i między miofibrylami i przebiega równolegle ze specjalizacją powierzchni kontaktowych komórek. Kardiomiocyty tworzą „koniec do końca”, „koniec do boku”, tworząc włókna mięśnia sercowego, i ogólnie tkanka jest strukturą podobną do sieci. Niektóre kardiomiocyty we wczesnych stadiach kardiomiogenezy są skurczowo-wydzielnicze. Następnie, w wyniku rozbieżnego różnicowania, powstają „ciemne” (kurczliwe) i „jasne” (przewodzące) miocyty, w których granulki wydzielnicze znikają, gdy pozostają w miocytach przedsionkowych. Tak powstaje różnica hormonalnych kardiomiocytów. Komórki te zawierają centralnie położone jądro z rozproszoną chromatyną, jedno lub dwa jądra. W cytoplazmie dobrze rozwinięty jest ziarnisty retikulum endoplazmatyczne, dyktiosomy kompleksu Golgiego, w ścisłym połączeniu z elementami, których są liczne granulki wydzielnicze o średnicy około 2 mikronów, zawierające materiał o dużej gęstości elektronowej. Następnie granulki wydzielnicze znajdują się pod sarcolemmą i są wydzielane do przestrzeni międzykomórkowej przez egzocytozę.
Ogólnie podczas histogenezy powstaje pięć rodzajów kardiomiocytów - praca (skurcz), zatok (rozrusznik serca), przejściowy, przewodzący i wydzielniczy. Pracujące (skurczowe) kardiomiocytytworzą ich łańcuchy (ryc. 9.10). To one, po skróceniu, zapewniają siłę skurczu całego mięśnia sercowego. Działające kardiomiocyty są w stanie
Ryc. 9.9Histogeneza tkanki mięśnia sercowego (schemat według P.P. Rumyantseva): ale- kardiomiocyty w ścianie rurki serca; b -kardiomiocyty w późnej embriogenezie; w- kardiomiocyty w okresie postanatalnym. 1 - kardiomiocyt; 2 - mitotycznie dzielący kardiomiocyt; 3 - miofilamenty i miofibryle
przesyłają sobie sygnały sterujące. Kardiomiocyty zatok (rozrusznik serca)potrafi automatycznie w określonym rytmie zmienić stan skurczu na stan rozluźnienia. Komórki odbierają sygnały kontrolne z włókien nerwowych, w odpowiedzi na które zmieniają rytm czynności skurczowej. Kardiomiocyty zatokowe (rozrusznik serca) przesyłają sygnały kontrolne przejściowe kardiomiocyty,a ten ostatni do przewodzących i pracujących kardiomiocytów. Prowadzenie kardiomiocytówtworzą łańcuchy komórek połączone ich końcami i znajdują się pod endo-
Ryc. 9.10Struktura tkanki mięśnia sercowego (mikrografia). Barwienie - żelazo hematoksylina:
1 - jądro kardiomiocytu; 2 - łańcuch kardiomiocytów; 3 - włóż koła
kard. Pierwsza komórka w łańcuchu odbiera sygnały kontrolne z kardiomiocytów zatokowych i przekazuje je do innych przewodzących kardiomiocytów. Komórki zamykające łańcuch przekazują sygnał przez przejściowe kardiomiocyty do pracowników. Kardiomiocyty wydzielniczepełnić specjalną funkcję. Wytwarzają hormon peptydowy kardiodilatynę, która krąży we krwi jak kardionatryna, powoduje zmniejszenie gładkich tętniczek miocytów, wzrost nerkowego przepływu krwi oraz przyspiesza filtrację kłębuszkową i wydalanie sodu. Wszystkie kardiomiocyty są pokryte błoną piwniczną.
Struktura kurczliwych (pracujących) kardiomiocytów.Komórki mają wydłużony (100–150 μm) kształt zbliżony do cylindrycznego. Ich końce są połączone ze sobą, dzięki czemu łańcuchy komórkowe tworzą tak zwane włókna funkcjonalne (o grubości do 20 mikronów). W obszarze styków komórki powstają tak zwane dyski wkładane (ryc. 9.10). Kardiomiocyty mogą rozgałęziać się i tworzyć sieć przestrzenną. Ich powierzchnie pokryte są membraną piwniczną, w którą wplecione są z zewnątrz włókna siatkowe i kolagenowe. Jądro kardiomiocytu (czasem dwa z nich) jest owalne i leży w środkowej części komórki. Na biegunach jądra koncentruje się niewiele organelli o ogólnym znaczeniu, z wyjątkiem agranularnej retikulum endoplazmatycznego i mitochondriów.
Specjalne organelle wywołujące skurcz nazywane są miofibrylami. Są słabo odizolowane od siebie, można je rozdzielić. Ich struktura jest podobna do struktury miofibryli miosymplastu włókna mięśni szkieletowych. Każdy mitochondria znajduje się w całym sarkomerie. Rurki T znajdujące się na poziomie linii Z są skierowane z powierzchni błony plazmatycznej na głębokość kardiomiocytu. Ich błony są połączone
w kontakcie z błonami gładkiej sieci endoplazmatycznej (sarkoplazmatycznej). Pętle tych ostatnich są wydłużone wzdłuż powierzchni miofibryli i mają boczne przedłużenia (układy L), które razem z trójnikami tworzą triadę lub diadę (ryc. 9.11, a). W cytoplazmie występują inkluzje glikogenu i lipidów, zwłaszcza wiele inkluzji mioglobiny. Mechanizm skurczu kardiomiocytów jest taki sam jak w przypadku miosymplastu.
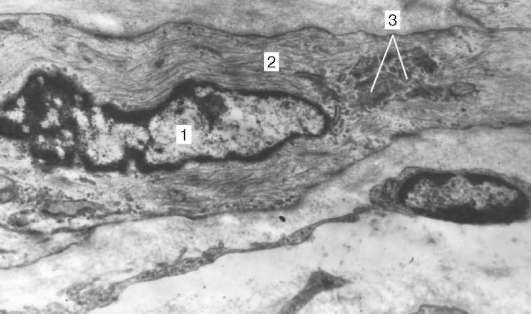
Organizacja kardiomiocytów w tkankę.Kardiomiocyty są połączone ze sobą przez rodzaj „od końca do końca”. Tworzą się tutaj dyski: te obszary wyglądają jak cienkie płytki o średnim powiększeniu mikroskopu świetlnego. W rzeczywistości końce kardiomiocytów mają nierówną powierzchnię, więc wypukłości jednej komórki wchodzą do koryta drugiej. Przekroje poprzeczne występów sąsiednich komórek są wzajemnie połączone interdigitacjami i desmosomami (ryc. 9.11, b).
Ryc. 9.11Struktura kardiomiocytów: ale- schemat (zgodnie z Yu. I. Afanasyev i V. L. Goryachkina); b- elektroniczna mikrografia dysku do wkładania. Wzrost o 20 000,1 - miofibryle; 2 - mitochondria; 3 - sieć sarkotubowa; 4 - trójniki; 5 - membrana piwnicy; 6 - lizosom; 7 - włóż dysk; 8 - desmosom; 9 - strefa przyłączenia miofibryli; 10 - styki szczelinowe; 11 - glikogen
Do każdego desmosomu od strony cytoplazmy zbliża się myofibryl la, który jest utrwalony do końca w kompleksie desmo-płytki. Tak więc, jednocześnie zmniejszając głód jednego kardiomiocytu, jest on przenoszony na inny. Boczne powierzchnie wypukłości kardiomiocytów są połączone przez nexusy (połączenia szczelinowe). Tworzy to między nimi wiązania metaboliczne i zapewnia synchronizację skurczów.
RegeneracjaW histogenezie tkanki mięśnia sercowego kambium nie występuje. Dlatego regeneracja tkanek przebiega na podstawie wewnątrzkomórkowych procesów hiperplastycznych. Jednocześnie proces poliploidii jest charakterystyczny dla kardiomiocytów ssaków, naczelnych i ludzi.
naród. Na przykład u małp nawet do 50% terminalnie zróżnicowanych kardiomiocytów w jądrze staje się tetra- i oktoploidalne. Kardiomiocyty poliploidalne występują z powodu mitozy akitokinetycznej, która prowadzi do wielonukleacji. W warunkach patologii układu sercowo-naczyniowego człowieka (reumatyzm, wrodzone wady serca, zawał mięśnia sercowego itp.) Regeneracja wewnątrzkomórkowa, poliploidalizacja jąder i pojawienie się wielojądrzastych kardiomiocytów odgrywają ważną rolę w kompensowaniu uszkodzenia kardiomiocytów.
9.3 GŁADKIE TKANINY MIĘŚNIOWE
Istnieją trzy grupy gładkich (niestriatowanych) tkanek mięśniowych (textus muscularis nonstriatus)oraz komórki: komórki mezenchymalne, nerwowe i mioepitelialne.
9.3.1 Tkanka mięśniowa pochodzenia mezenchymalnego
Histogeneza.Ta tkanka dzieli się na dwa typy: trzewny i naczyniowy. W zarodkowej histogenezie nawet mikroskopia elektronowa jest trudna do rozróżnienia między mezenchymalnymi progenitorami fibroblastów i gładkimi miocytami. W słabo zróżnicowanych gładkich miocytach rozwija się ziarnisty retikulum endoplazmatyczne, kompleks Golgiego. Cienkie włókna są zorientowane wzdłuż długiej osi komórki. W miarę ich rozwoju zwiększa się rozmiar komórki i liczba filamentów w cytoplazmie. Stopniowo zwiększa się objętość cytoplazmy zajmowanej przez włókna kurczliwe, ich lokalizacja w cytoplazmie staje się coraz bardziej uporządkowana. Aktywność proliferacyjna gładkich miocytów w miogenezie jest stopniowo zmniejszana. Dzieje się tak w wyniku wzrostu czasu trwania cyklu komórkowego, wyjścia komórek z cyklu reprodukcyjnego i przejścia do zróżnicowanego stanu. Różnicując, syntetyzują składniki macierzy międzykomórkowej, kolagen błony podstawnej, a także elastynę. W ostatecznych komórkach (miocytach) zdolność syntezy jest zmniejszona, ale nie zanika całkowicie.
Struktura i funkcjonowanie komórek.Gładki miocyt to komórka w kształcie wrzeciona o długości 20-500 mikronów i szerokości 5-8 mikronów. Rdzeń ma kształt pręta, umieszczony w jego środkowej części. Kiedy miocyt kurczy się, jego rdzeń wygina się, a nawet skręca (ryc. 9.12–9.14).
Struktura definitywnie gładkich miocytów (leiomiocytów), które tworzą narządy wewnętrzne i ściany naczyń krwionośnych, ma wiele cech wspólnych, ale jednocześnie charakteryzuje się heteromorfizmem. Tak więc w ścianach żył i tętnic znajdują się jajowate, wrzecionowate, przetwarzają miocyty o długości 10-40 mikronów, czasem dochodzące nawet do 140 mikronów. Najdłuższa gładka mio-
cytaty docierają do ściany macicy - do 500 mikronów. Średnica miocytów wynosi od 2 do 20 mikronów. W zależności od charakteru wewnątrzkomórkowych procesów biosyntezy rozróżnia się miocyty skurczowe i wydzielnicze. Te pierwsze specjalizują się w funkcji skurczu, ale jednocześnie zachowują aktywność wydzielniczą.
Wydzielające miocyty w swojej ultrastruktury przypominają fibroblasty, ale zawierają w swoich cytoplazmie wiązki cienkich włókien mięśniowych zlokalizowanych na obrzeżach komórki. Kompleks Golgiego, ziarnisty retikulum endoplazmatyczne, wiele mitochondriów, granul glikogenu, wolne rybosomy i polisomy są dobrze rozwinięte w cytoplazmie. Ze względu na stopień dojrzałości takie komórki są klasyfikowane jako słabo zróżnicowane. Włókna aktynowe w cytoplazmie tworzą trójwymiarową sieć, wydłużoną głównie wzdłużnie. Końce filamentów są łączone razem i za pomocą plazmolematu za pomocą specjalnych białek sieciujących. Obszary te są wyraźnie widoczne na mikrografach elektronowych jako gęste ciała. Monomery miozyny znajdują się obok włókien aktynowych. Plasmolemma tworzy wtargnięcia - jaskinie, w których skoncentrowane są jony wapnia. Sygnał skurczu zwykle pochodzi z włókien nerwowych. Mediator, który wyróżnia się na tle swoich warunków, zmienia przepuszczalność plazmy. Uwalniane są jony wapnia, co pociąga za sobą zarówno polimeryzację miozyny, jak i interakcję miozyny z aktyną.
Myofilamenty aktyny są wciągane pomiędzy
Ryc. 9.12Struktura gładkiego miocytu (schemat):
a) w- podczas relaksu; b d- przy największej redukcji; g- z niepełną redukcją; w- powiększone obrazy odcinków otoczonych fragmentami alei b. 1 - plazmolemma; 2 - gęste małe ciała; 3 - rdzeń; 4 - endoplazma; 5 - kompleksy skurczowe; 6 - mitochondria; 7 - membrana piwnicy; 8 - cienkie miofilamenty aktynowe; 9 - miozyna (gruba) miofilamenty
Ryc. 9.13Ultrastruktura różnicowania gładkiego miocytu w ścianie oskrzeli:
1 - rdzeń; 2 - cytoplazma z miofilamentami; 3 - Kompleks Golgiego, wzrost o 35 000 (lek A. L. Zashikhina)
spotykają się nowe, gęste plamy, siła jest przenoszona na plazmolemmę, a cała komórka ulega skróceniu (patrz ryc. 9.12). Kiedy zanikają sygnały z układu nerwowego, jony wapnia przemieszczają się z cytoplazmy do jaskini i kanalików retikulum endoplazmatycznego, miozyna ulega depolimeryzacji, a miofibryle rozpadają się. Skurcz ustaje. Tak więc kompleksy aktynomomiozyny występują w gładkich miocytach tylko w okresie skurczu w obecności wolnych jonów wapnia w cytoplazmie.
Miocyty są otoczone membraną piwnicy. W niektórych obszarach powstają w nim „okna”, więc plazmolemma sąsiednich miocytów zbiega się. Tutaj powstają ogniwa i między komórkami powstają nie tylko wiązania mechaniczne, ale także metaboliczne. Na „osłonach” błony podstawnej między miocytami znajdują się włókna elastyczne i siatkowe, łączące komórki w pojedynczy kompleks tkanek. Gładkie miocyty syntetyzują proteoglikany, glikoproteiny, prokolagen, proelastynę, z których powstają włókna kolagenowe i elastyczne oraz amorficzny składnik macierzy międzykomórkowej. Oddziaływanie miocytów odbywa się za pomocą mostków cytoplazmatycznych, wzajemnych wypukłości, nexusów, desmosomów, odcinków styków błonowych powierzchni miocytów.
RegeneracjaGładka tkanka mięśniowa gatunków trzewnych i naczyniowych ma znaczną wrażliwość na działanie czynników ekstremalnych. W aktywowanych miocytach wzrasta poziom procesów biosyntetycznych, których morfologiczną ekspresją jest synteza białek kurczliwych, powiększenie i hiperchromatoza jądra, przerost jąder, wzrost wskaźnika cytoplazmatycznego jądra, wzrost liczby wolnych rybosomów i polisomów oraz

Ryc. 9.14Struktura tkanki mięśni gładkich (schemat wolumetryczny) (według R.V. Krstic, ze zmianami):
1 - wrzecionowate gładkie miocyty; 2 - cytoplazma miocytów; 3 - jądra miocytów; 4 - plazmolemma; 5 - membrana piwnicy; 6 - powierzchowne pęcherzyki pinocytotyczne; 7 - połączenia międzykomórkowe; 8 - zakończenie nerwowe; 9 - włókienka kolagenowe; 10 - mikrofilamenty
enzymy, tlenowa i beztlenowa fosforylacja, transport błon. Regeneracja komórek odbywa się zarówno ze względu na zróżnicowane komórki ze zdolnością do wejścia w cykl mitotyczny, jak i ze względu na aktywację elementów kambialnych (miocytów o małej objętości). Pod działaniem wielu szkodliwych czynników obserwuje się fenotypową transformację kurczliwych miocytów w wydzielniczą. Ta transformacja jest często obserwowana z uszkodzeniem błony naczyniowej, tworzeniem się rozrostu błony wewnętrznej wraz z rozwojem miażdżycy.
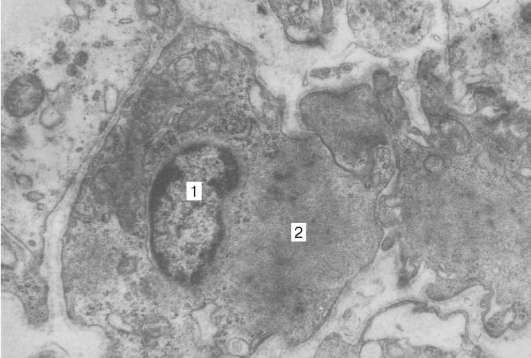
Ryc. 9.15Ultrastruktura miopigmentocytów (lek N. N. Sarbaeva): 1 - jądro; 2 - włókna mięśniowe, wzrost o 6000
9.3.2 Mezenchymalna tkanka mięśniowa w narządach
Miocyty są łączone w wiązki, między którymi znajdują się cienkie warstwy tkanki łącznej. Siatkowe i elastyczne włókna otaczające miocyty są wplecione w te warstwy. Naczynia krwionośne i włókna nerwowe przechodzą przez warstwy pośrednie. Końce tych ostatnich nie kończą się bezpośrednio na miocytach, ale między nimi. Dlatego po nadejściu impulsu nerwowego mediator rozprzestrzenia się rozproszony, pobudzając wiele komórek naraz. Gładka tkanka mięśniowa pochodzenia mezenchymalnego jest reprezentowana głównie w ścianach naczyń krwionośnych i wielu pustych narządach wewnętrznych.
Gładka tkanka mięśniowa w określonych narządach ma nierówne właściwości funkcjonalne. Wynika to z faktu, że na powierzchni narządów znajdują się różne receptory dla określonych substancji biologicznie czynnych. Dlatego w przypadku wielu leków ich reakcja nie jest taka sama. Możliwe jest, że różne właściwości funkcjonalne tkanek są związane ze specyficzną organizacją molekularną włókien aktyny.
9.3.3 Tkanka mięśniowa pochodzenia nerwowego
Tkanka mięśniowa tęczówki i ciała rzęskowego jest czwartym rodzajem tkanki kurczliwej. Miocyty tej tkanki rozwijają się z komórek podstawy nerwowej jako części wewnętrznej ściany kielicha wzrokowego. Z rzędu
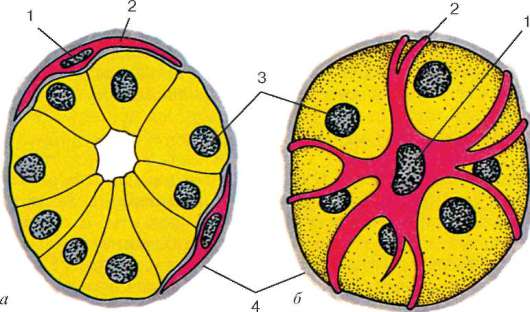
Ryc. 9.16Komórki mioepitelialne w końcowym odcinku gruczołu ślinowego (schemat według G. S. Katinas):
ale- przekrój; b- widok z powierzchni. 1 - jądra komórek mioepitelialnych; 2 - procesy komórek mioepitelialnych; 3 - jądra wydzielniczych komórek nabłonkowych; 4 - membrana piwnicy
elementy mięśnia kręgowego tęczówki wykazują zróżnicowane zróżnicowane zróżnicowanie. Tak więc tkankę mięśniową u gadów i ptaków reprezentują prążkowane włókna wielojądrzaste, które są bardzo podobne do mięśni szkieletowych. U ssaków i ludzi główną strukturalną i funkcjonalną jednostką mięśni tęczówki jest gładki jednojądrzasty miocyt lub miopigmentocyt. Te ostatnie mają pigmentowany korpus zawierający jeden rdzeń, wyjęty poza część skurczową w kształcie wrzeciona (ryc. 9.15).
Cytoplazma komórek zawiera dużą liczbę mitochondriów i granulek pigmentowych, które pod względem wielkości i kształtu są podobne do granulek nabłonka pigmentowego. Miofilamenty w miopigmentocytach są podzielone na cienkie (7 nm) i grube (1,5 nm), pod względem wielkości i lokalizacji przypominają miofilamenty gładkich miocytów. Każdy miopigmentocyt jest otoczony błoną piwnicy. W pobliżu procesów cytoplazmatycznych miocytów znaleziono włókna nerwowe wolne od mieliny. W zależności od kierunku procesów (prostopadłego lub równoległego do krawędzi źrenicy) miocyty tworzą dwa mięśnie - zwężającą się i rozszerzającą źrenicę.
RegeneracjaKilka badań wykazało niską aktywność regeneracyjną po uszkodzeniu lub jego braku.
9.3.4 Naskórkowe komórki mięśniowe
Komórki mioepitelialne rozwijają się z zarodka naskórka. Występują w gruczołach potowych, sutkowych, ślinowych i łzowych i mają wspólnych poprzedników z komórkami wydzielniczymi. Mioepitelialny
komórki przylegają bezpośrednio do odpowiedniego nabłonka i mają ze sobą wspólną błonę podstawną. Podczas regeneracji te i inne komórki są również przywracane ze zwykłych nisko zróżnicowanych prekursorów. Większość komórek mioepitelialnych ma kształt gwiazdy. Komórki te są często nazywane koszem: ich procesy obejmują odcinki końcowe i małe kanały gruczołów (ryc. 9.16). Rdzeń i organelle o ogólnym znaczeniu znajdują się w ciele komórki, aw procesach - aparacie kurczliwym, zorganizowanym jak w komórkach tkanki mięśniowej typu mezenchymalnego.
Pytania bezpieczeństwa
1. Klasyfikacja genetyczna tkanki mięśniowej. Jednostki strukturalne i funkcjonalne różnych rodzajów tkanki mięśniowej.
2. Prążkowana tkanka mięśni szkieletowych: rozwój, budowa, morfologiczne podstawy skurczu mięśni. Regeneracja
3. Prążkowana tkanka mięśnia sercowego: rozwój, specyficzna struktura różnych rodzajów kardiomiocytów, regeneracja.
4. Odmiany gładkich miocytów: źródła rozwoju, topografia w ciele, regeneracja.
Histologia, embriologia, cytologia: podręcznik / Yu. I. Afanasyev, N. A. Yurina, E. F. Kotovsky i inni. - 6 edycja, Rev. i dodaj. - 2012 r. - 800 s. : chory
Mięśnie ludzkiego ciała są tworzone głównie przez tkankę mięśniową, składającą się z komórek mięśniowych. Rozróżnij gładką i prążkowaną tkankę mięśniową. Gładka tkanka mięśniowa buduje muskulaturę, która jest częścią niektórych narządów wewnętrznych, i prążkowanytworzy mięsień szkieletowy. Wspólną właściwością tkanki mięśniowej jest jej pobudliwość, przewodnictwo i kurczliwość.
Prążkowana tkanka mięśniowa różni się od gładkiej, wyższej pobudliwości, przewodności i kurczliwości. Komórki mięśnia prążkowanego mają bardzo małą średnicę i dużą długość (do 10-12 cm). W związku z tym są one nazywane włókna.
Podobnie jak inne komórki, komórki mięśniowe mają tzw. Protoplazmy sarkoplazma(z greckiego. Sarcos - mięso). Błona komórkowa mięśni zwana sarcolemma. Wewnątrz włókna mięśniowego znajduje się wiele jąder i innych składników komórek.
Skład włókien mięśniowych obejmuje dużą liczbę jeszcze cieńszych włókien - miofibryle, które z kolei składają się z najcieńszych nici - protofibryli. Protofibryle - to jest aparat kurczliwy komórki mięśniowej, są to specjalne białka kurczliwe - miozyna i aktyna. Mechanizm skurcze mięśni Jest to złożony proces przemian fizycznych i chemicznych we włóknach mięśniowych z obowiązkowym udziałem aparatu kurczliwego. Uruchomienie tego mechanizmu odbywa się za pomocą impulsu nerwowego, a energię do procesu redukcji dostarcza adenozyny kwas trifosforowy (ATP). Pod tym względem cechą struktury włókien mięśniowych jest również duża liczba mitochondriów, które zapewniają włóknom mięśniowym niezbędną energię. Rozluźnienie włókna mięśniowego, zdaniem wielu naukowców, odbywa się pasywnie, ze względu na elastyczność sarcolemmy i domięśniowej tkanki łącznej.
Struktura, kształt i klasyfikacja mięśni szkieletowych
Jednostką anatomiczną najbardziej aktywnej części układu mięśniowego człowieka - mięśni szkieletowych lub prążkowanych - jest mięsień szkieletowy. Mięsień szkieletowy jest organem utworzonym przez prążkowaną tkankę mięśniową, która zawiera również tkankę łączną, nerwy i naczynia krwionośne.
Każdy mięsień jest otoczony swoistym „przypadkiem” tkanki łącznej - powięź.
W struktura zewnętrzna mięśnie rozróżniają głowę ścięgna odpowiadającą początkowi mięśnia, brzuchowi mięśnia lub ciału utworzonemu z włókien mięśniowych oraz koniec ścięgna mięśnia lub ogona, którym mięsień przyczepia się do innej kości. Zazwyczaj ogon mięśnia jest ruchomym punktem przywiązania, a początek jest ustalony. W trakcie ruchu ich funkcje mogą się zmieniać: ruchome punkty stają się nieruchome i odwrotnie.
Kształt mięśni jest bardzo zróżnicowany i w dużej mierze zależy od funkcjonalnego celu mięśnia. Rozróżnij mięśnie długie, krótkie, szerokie, okrągłe i inne. Długo mięśnie znajdują się na kończynach, krótki - tam, gdzie ruch jest niewielki (na przykład między kręgami). Szerokie mięśnie znajdują się głównie na ciele (mięśnie brzucha, pleców, klatki piersiowej). Okrągłe mięśnie są rozmieszczone wokół otworów ciała i zwężają je po skurczeniu. Te mięśnie są nazywane zwieracze. Jeśli mięsień ma jedną głowę, nazywa się go prostym, jeśli dwa lub więcej - złożonym (na przykład biceps, triceps i czworogłowy). Ważną cechą morfologiczną jest lokalizacja włókien mięśniowych. Rozróżnij równoległy, ukośny, poprzeczny i okrągły układ włókien (w zwieraczach). jeśli z ukośnym układem włókien mięśniowych łączą się tylko z jednej strony ścięgnami, wówczas nazywane są mięśnie pierzastyjeśli po obu stronach - rozwidlony.
Funkcjonalnie można podzielić mięśnie zginacze i prostowniki, rotatory na zewnątrz i rotatory wewnątrz, mięśnie przywodziciela i absolutorium. Mięśnie są również wydzielane - synergetyki i mięśnie - antagoniści. Te pierwsze tworzą grupę mięśni, które przyjaźnie wykonują każdy ruch, skurcz tego drugiego powoduje ruchy przeciwne.
Główne grupy mięśni
Mięśnie szkieletowe osoby dorosłej stanowią około 40% jego masy ciała. U noworodków i małych dzieci mięśnie stanowią nie więcej niż 20–25% masy ciała, zaś w starszym wieku następuje stopniowy spadek masy mięśniowej do 25–30% masy ciała. W sumie w ludzkim ciele znajduje się ponad 600 mięśni.
Według lokalizacji, tj. na podstawie topograficznej rozróżnia się mięśnie pleców, klatki piersiowej, brzucha, głowy, szyi, kończyn górnych i dolnych.
Mięśnie pleców są podzielone na powierzchowne i głęboki. To powierzchowne mięśnie do pleców należą: mięsień trapezowy, latissimus dorsi, mięśnie romboidalne (większe i mniejsze), mięśnie zębate (górne i dolne).
Mięśnie te biorą udział w ruchach kończyn górnych i zmianach objętości. skrzynia (akt oddychania).
Mięśnie głębokich pleców są reprezentowane przez liczne mięśnie umieszczone wzdłuż kręgosłupa. Są one przywiązane głównie do procesów kręgów i biorą udział w ruchach kręgosłupa z powrotem i na boki.
Mięśnie klatki piersiowej są podzielone na mięśnie związane z obręczą barkową i kończyna górna (mięśnie piersiowe większe i mniejsze, mięśnie podobojczykowe i przednie zębate) , i własne mięśnie klatki piersiowej (zewnętrzne wewnętrzne mięśnie międzyżebrowe). Wprawiają w ruch i wzmacniają kończyny górne na ciele oraz uczestniczą w funkcji oddychania. Przepona należy również do mięśni oddechowych (oddziela jamę klatki piersiowej od jamy brzusznej).
Mięśnie brzucha są reprezentowane przez zewnętrzne i wewnętrzne mięśnie skośne, poprzeczne i proste brzucha, a także kwadratowy mięsień dolnej części pleców.
Mięśnie brzucha odbytnicy uczestniczą w zginaniu ciała do przodu, a mięśnie skośne zapewniają nachylenie kręgosłupa w bok i jego obroty wraz z klatką piersiową w prawo i w lewo. Mięśnie brzucha uczestniczą nie tylko w ruchach tułowia i klatki piersiowej. Tworzą ścianę jama brzuszna i ze względu na ich ton utrzymują narządy wewnętrzne w normalnej pozycji. Zmniejszając je, zwiększają ciśnienie w jamie brzusznej, tworząc tzw brzuszny. Mięśnie brzucha ułatwiają wypróżnienia (wypróżnienia), oddawanie moczu oraz wydalenie płodu u kobiet podczas porodu. Skurcz mięśni brzucha przyczynia się również do przepływu krwi w układzie żylnym i wykonywania ruchów oddechowych, ale także do powstawania przepukliny. W przypadku przepukliny dochodzi do wysunięcia narządów wewnętrznych (jelit, żołądka itp.) Z jamy brzusznej pod skórą.
Wśród mięśni głowy wyróżnia się mięśnie do żucia i mięśnie twarzy. To mięśnie do żucia stosuje czasowe, żucie, pterygoid. Skurcz tych mięśni powoduje złożone ruchy żucia. żuchwa. Mięśnie naśladujące różnią się od innych mięśni ludzkiego ciała tym, że są przymocowane do skóry twarzy jednym, a czasem dwoma, końcami. Skurcz tych mięśni to przemieszczenie skóry, tworzenie się fałd i zmarszczek. To definiuje mimikę twarzy, tj. taki czy inny wyraz twarzy. Manifestacja złożonych doznań (emocji) - radości, wstydu, pogardy, żalu i bólu itp. - zdeterminowany licznymi kombinacjami skurczów mięśni twarzy, podporządkowanych impulsom dochodzącym do nich z kory mózgowej wzdłuż nerwu twarzowego. Okrągłe mięśnie oka i ust są również mięśniami imitującymi. Mięśnie szyi przechylają głowę, przechylają ją i obracają. Mięśnie przyczepione do kości gnykowej podczas skurczu zmieniają położenie języka i krtani podczas połykania i wydawania dźwięków.
Mięśnie kończyny górnej są podzielone na mięśnie pas barkowy i mięśnie wolnej kończyny górnej. Mięśnie obręczy barkowej otaczają staw barkowy, zapewniając w nim liczne ruchy. Najważniejszym z nich jest mięsień naramienny. Podczas skurczu mięsień ten zgina ramię staw barkowy i podnosi rękę do pozycji poziomej.
Mięśnie wolnej kończyny górnej obejmują mięśnie barku, przedramienia i dłoni.
W przedniej części ramienia znajduje się grupa mięśni - zginaczepowrót - prostowniki. Wśród mięśni przedniej grupy najważniejszy jest mięsień bicepsowy barku (biceps), a plecy, mięsień trójgłowy barku.
Mięśnie przedramienia są również reprezentowane na przedniej powierzchni przez zginacze, a na plecach przez prostowniki. Mięśnie dłoni znajdują się tylko na powierzchni dłoniowej. Wśród nich wyróżnia się - długi mięsień dłoniowy i zginacze palców. Ze względu na obecność własnego układu mięśniowego, szczególnie palców kciuk, uzyskują większą mobilność i są zdolne do różnych ruchów, w tym maksymalnego zginania i rozciągania, co jest niezwykle ważne podczas pracy.
Mięśnie znajdujące się w okolicy pasa kończyny dolnej poruszają nogę staw biodrowy, a także ze stałą kończyną, zginają kręgosłup w okolicy lędźwiowej. Przednia grupa mięśni obejmuje jeden główny mięsień - mięśnie biodrowe. Wśród tylnej grupy mięśni obręczy miednicy - duże, średnie i małe mięśnie pośladkowe.
Kończyny dolne mają bardziej masywny szkielet niż górne; ich mięśnie mają wielką siłę, ale jednocześnie mniej różnorodności i ograniczony zakres ruchu.
Na udzie z przodu znajduje się najdłuższy mięsień krawiecki w ciele człowieka (do 50 cm). Zagina nogę w stawach biodrowych i kolanowych.
Mięsień czworogłowy kości udowej leży głębiej niż mięsień krawiecki, otaczając kość udową niemal ze wszystkich stron.
Główną funkcją tego mięśnia jest przedłużenie stawu kolanowego. W pozycji stojącej mięsień czworogłowy nie daje staw kolanowy zginać się.
Na stopie rozróżnia się mięśnie pleców i podeszwowe. Mięśnie te biorą udział w zgięciu i wyprostowaniu palców, a także w ich zbieżności i hodowli.