Quante vertebre sacrali ha una lucertola. Sistema muscolo-scheletrico di rettili caratteristica generale dello scheletro
ARGOMENTO 14. SCHELETRO DI UN REBUMAN
POSIZIONE SISTEMATICA DELL'OGGETTO
Sottotipo Vertebrata, Vertebrata
Reptile Class, Reptilia
1. Squad Scale, Squamata
Rappresentante - Varan Grey, Varanus griseus Daud.
2. Coccodrilli rosa, Crocodilia
Rappresentante - Alligatore del Mississippi, Alligator mississippiensts Daud.
3. Turtle Squad, Chelonia (Testudines)
Rappresentante - Tartaruga palustre, Etnys orbicular is L.
MATERIALE E ATTREZZATURA
Scheletri montati di lucertole monitor, serpenti, tartarughe - uno per gruppo di studenti; teschi di coccodrillo (o manichini) - 1-2 per gruppo. Per uno o due studenti sono richiesti:
1. Scheletro pieghevole di una lucertola monitor.
2. Il cranio della tartaruga.
3. Aghi da dissezione - 2.
JOB
Considera lo scheletro di una lucertola monitor nel suo insieme. Trova le sue parti principali: teschio, colonna vertebrale, scheletro di arti accoppiati e cinture. Considera le caratteristiche strutturali dello scheletro di un serpente e una tartaruga. Considera i dettagli della struttura dello scheletro di una lucertola monitor e dei teschi di una lucertola monitor, coccodrillo e tartaruga. Confronta la struttura del cranio di un coccodrillo, controlla la lucertola, il serpente e la tartaruga.
Fai i seguenti disegni:
1. Il cranio della lucertola monitor sul lato.
2. Il cranio della lucertola monitor dal basso.
4. La zampa anteriore di una lucertola monitor.
6. L'arto posteriore della lucertola monitor.
7. Cranio di coccodrillo in cima.
8. Il teschio di coccodrillo dal basso.
9. Il cranio della tartaruga sul lato.
DESCRIZIONE DELLO SCHELETRO
Scheletro assiale. La differenziazione dello scheletro assiale, o colonna vertebrale, in dipartimenti è più pronunciata nei rettili che negli anfibi. La colonna cervicale (pars cervicalis) è sempre composta da diverse vertebre, di cui le due anteriori hanno una struttura speciale. La prima vertebra cervicale è chiamata atlante o atlante (atlante; Fig. 81, A). È privo del corpo vertebrale e ha la forma di un anello diviso in due parti. Sulla superficie frontale inferiore di questa vertebra è presente una cavità articolare, collegata in modo mobile al condilo del cranio (vedi sotto). La seconda vertebra cervicale - epistrophy (epistrophaeus; Fig. 81, B), ha di fronte un grande processo simile a un dente (processus odontoideus, Fig. 81, 1), che è il corpo della prima vertebra cervicale fusa con epistrofia. Il processo simile a un dente entra liberamente nell'apertura inferiore dell'atlante. Questa struttura delle prime vertebre cervicali offre una maggiore mobilità della testa. Le rimanenti vertebre cervicali hanno il solito dispositivo (vedi sotto); molti hanno costole a collo corto.
Fig. 81. Vertebre vertebrali
A - atlante; B - un'epistrofia; B - vertebra toracica; G - sezione longitudinale della vertebra toracica:
1 - processo simile a un dente dell'epistrofia, 2 - corpo vertebrale, 3 - arco superiore, 4 - processo spinoso, 5 - canale per il midollo spinale, 6 - processo articolare anteriore, 7 - processo articolare posteriore
Le divisioni toracica e lombare non sono del tutto distinte e sono generalmente considerate come una singola divisione (pars thoracolumbalis). In realtà, la regione toracica è la parte della colonna vertebrale in cui le costole che si estendono dalle vertebre con l'estremità inferiore si attaccano allo sterno. Le vertebre della regione lombare portano costole che non raggiungono lo sterno. I corpi vertebrali (Fig. 81, 2) sono concavi davanti e convessi dietro; tali vertebre sono chiamate percentuale. Sopra il corpo vertebrale si alzano gli archi superiori (arcus neuralis; Fig. 81, 3), che termina con un processo spinoso (processusspinosus; Fig. 81, 4). Nel canale formato dagli archi superiori, si trova midollo spinale.
I processi anteriori (processus articularis anterior; Fig. 81, 6) e posteriori (processus articularis posteriore; Fig. 81, 7) i processi articolari partono dalle parti anteriore e posteriore della base degli archi superiori. Questi processi accoppiati si collegano ai processi articolari delle vertebre vicine e contribuiscono a una maggiore forza della colonna vertebrale durante la flessione. Ai lati del corpo vertebrale (vicino alla base degli archi superiori) ci sono piccole rientranze a cui si attaccano le costole.
La sezione sacrale (pars sacralis) è costituita da due vertebre, che sono caratterizzate da processi trasversali fortemente sviluppati (processus transversus); sono uniti dalle ossa del bacino. La sezione della coda (pars caudalis) è rappresentata da numerose vertebre, che diminuiscono gradualmente di dimensioni.
Questa struttura della colonna vertebrale è tipica della classe dei rettili, ma in alcuni gruppi subisce cambiamenti secondari. In particolare, nei serpenti, in relazione alla riduzione degli arti accoppiati e all'emergere di un diverso tipo di movimento - strisciare sull'addome piegando il tronco - la colonna vertebrale è chiaramente divisa solo nelle sezioni del tronco e della coda. Tutte le vertebre del tronco hanno costole mobili, le cui estremità inferiori sono libere (lo sterno dei serpenti è assente) e appoggiano contro le cicatrici addominali cornee.

Fig. 82. Scheletro di una tartaruga di palude
A - carapace; B - plastron:
1 - sezione del tronco della colonna vertebrale, 2 - placche costali, 3 - placche marginali, 4 - coracoide, 5 - scapola, 6 - ilio7 - osso pubico; 8 - osso ischiatico
Nelle tartarughe, lo scheletro assiale prende parte alla formazione della base ossea del loro carapace. Lo scudo superiore del guscio - carapace (caparax; Fig. 82, A) - è composto da più file di placche ossee. La fila centrale (non accoppiata) di queste placche è formata dalla fusione di processi spinosi e trasversali estesi e appiattiti delle vertebre del tronco con ossa della pelle; ai lati della fila centrale sono allineate file di placche ossee fuse con nervature allargate. Il bordo del carapace è formato da placche ossee di origine tegumentaria. Pertanto, la colonna vertebrale del busto è fissa e saldamente fusa con lo scudo dorsale del carapace. La colonna cervicale e caudale sono mobili. Allo stesso tempo, le vertebre cervicali anteriori sono opistocellulari (il corpo vertebrale è convesso nella parte anteriore e concavo nella parte posteriore), quelli posteriori sono longitudinali e tra questi due gruppi c'è una vertebra il cui corpo ha una superficie convessa sia davanti che dietro.
Skull. Rispetto agli anfibi, il cranio del rettile è caratterizzato da un'ossificazione molto più completa. Una certa quantità di cartilagine viene conservata solo nella capsula olfattiva e nella regione uditiva. Le parti assiali e viscerali del cranio sono poste separatamente nell'embrione, ma negli animali adulti rappresentano un'unica entità. La composizione del cranio comprende sia ossa della cartilagine (sostitutive o primarie) sia numerose ossa della pelle (tegumentarie o secondarie). È conveniente usare il cranio di una grande lucertola, la lucertola monitor, come oggetto principale di studio.

Fig. 83. Cranio della lucertola monitor
A è sul lato; B: dal basso; B: dall'alto; G - indietro:
1 - osso occipitale principale, 2 - osso occipitale laterale, 3 - osso occipitale superiore, 4 - forame occipitale grande, 5 - condilo occipitale, 6 - osso anteriore, 7 - principale sfenoide, 8 - apri, 9 - osso parietale, 10 - osso frontale, 11 - osso nasale, 12 - osso prefrontale, 13 - osso preorbitale, 14 - osso lacrimale, 15 - fossa temporale superiore, 16 - osso frontale posteriore, 17 - osso squamoso , 18 - l'osso mascellare, 19 - l'osso mascellare, 20 - l'osso zigomatico, 21 - la rottura dell'arco temporale inferiore a causa della riduzione dell'osso quadrato zigomatico, 22 - osso quadrato, 23 - osso pterigoideo, 24 - osso palatino, 25 - osso sopra-pterigoideo, 26 - osso trasversale, 27 - osso arcuato, 28 - osso del dente, 29 - osso angolare, 30 - osso articolare, 31 - osso coronarico
Cranio assiale Nella regione occipitale del cranio ci sono tutte e quattro le ossa occipitali: l'occipitale principale (basioccipitale, Fig. 83, 1), due occipitali laterali (occipitale laterale; Fig. 83, 2) e occipitale superiore (sopra occipitale; Fig. 83, 3). Queste ossa di origine primaria circondano il grande forame occipitale (forame occipitale magnum; Fig. 83, 4). Le ossa occipitali inferiori e laterali insieme formano l'unico condilo occipitale (a differenza degli anfibi) (condylus occipitalis; Fig. 83, 5), articolato con la prima vertebra cervicale - l'atlante. L'articolazione della testa con il collo usando solo un condilo, in combinazione con le caratteristiche strutturali delle prime due vertebre cervicali già considerate, conferisce alla testa del rettile una mobilità significativa.
Nella regione uditiva delle ossa della cartilagine, solo l'osso anteriore accoppiato (prooticum; Fig. 83, 6) rimane indipendente, mentre l'orecchio superiore si fonde con l'osso occipitale superiore e l'orecchio posteriore con l'occipitale laterale.
Il setto interorbitale dei rettili è sottile, membranoso, e solo coccodrilli e lucertole hanno piccole ossificazioni separate, apparentemente corrispondenti alle ossa a forma di cuneo. La capsula olfattiva non ha ossificazione.
Alla base del cranio, di fronte all'osso occipitale principale, c'è un osso sfenoide principale tegumentario piuttosto grande (basefenoideum; Fig. 83, 7). Il suo processo stretto anteriore è omologa al parasfenoide (parasphenoideum), che è notevolmente ridotto nei rettili. Nella parte anteriore della parte inferiore del cranio, sotto la sezione olfattiva, c'è un apriscatole accoppiato (vomer; Fig. 83, 8), anch'esso di origine tegumentaria.
Il tetto del cranio è rappresentato da numerose ossa tegumentarie, alcune delle quali cadono e ricoprono il cranio dai lati. Questi includono le ossa parietale (parietale; Fig. 83, 9), frontale (frontale; Fig. 83, 10) e nasale (nasale; Fig. 83, 11). Di fronte alle ossa frontali, le ossa prefrontali (praefrontale; Fig. 83, 12) e preorbitale (praeorbitale; Fig. 83, 13) accoppiate di solito si trovano e sotto di esse nella parete anteriore dell'orbita sono accoppiati lacrimali (lacrimale; Fig. 83, 14) ).
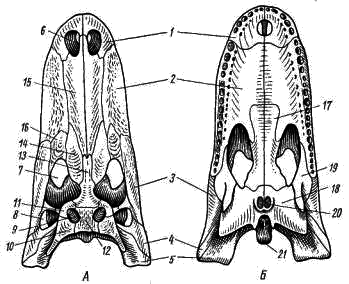
Fig. 84. Crocodile Skull (Mississippi Alligator).
A: dall'alto; B - fondo:
1 - osso mascellare, 2 - osso mascellare, 3 - osso zigomatico, 4 - osso zigomatico quadrato, 5 - osso quadrato, 6 - narice esterna, 7 - orbita, fossa temporale 8 laterale, 9 - fossa temporale superiore, 10 - osso squamoso, 11 - l'osso frontale (postorbitale), 12 - l'osso parietale, 13 - l'osso frontale, 14 - l'osso prefrontale, 15 - l'osso nasale, 16 - l'osso lacrimale, 17 - l'osso palatino, 18 - l'osso pterigoideo, 19 - l'osso trasversale , 20 - cigni (aperture interne delle narici), 21 - condilo occipitale
Delle rimanenti ossa tegumentarie del cranio assiale, le ossa che prendono parte alla formazione dei cosiddetti archi temporali sono di particolare interesse. Il coccodrillo nel tetto del cranio all'esterno dell'osso parietale (Fig. 84, 12) ha un foro su ciascun lato - la fossa temporale superiore (Fig. 84, 9). Sul bordo esterno, la fossa temporale superiore è delimitata dalle ossa posteriori frontali o postorbitali (postfrontale-postorbitale, Fig. 84, 11) e squamose (squamosio; Fig. 84, 10). Queste due ossa insieme formano l'arco temporale superiore. Sul lato del cranio dietro l'orbita sono presenti fosse temporali laterali (Fig. 84, 8), delimitate esternamente dagli archi temporali inferiori. Ogni arco temporale inferiore è composto da due ossa: lo zigomatico (jugale; Fig. 84, 3) e il quadrato-zigomatico (quadrato-jugale; Fig. 84, 4). L'arco temporale inferiore si collega alla mascella superiore: l'osso zigomatico cresce al mascellare e l'osso quadrato-zigomatico cresce al quadrato. Questo tipo di cranio, come un coccodrillo - con due fosse temporali e due archi temporali, è chiamato diapside (a due braccia).
In una lucertola monitor, la fossa temporale superiore (Fig. 83, 15) è delimitata dall'arco temporale superiore completo (postfrontale - squamosum; Fig. 83, 16-17). Come parte dell'arco temporale inferiore, l'osso quadrato-zigomatico è stato ridotto e solo l'osso zigomatico è stato preservato (Fig. 83 , 20); le fosse temporali laterali non sono quindi aperte dall'esterno e rimangono aperte. Pertanto, il cranio della lucertola monitor può essere considerato come un cranio di tipo diapside, ma con un arco inferiore ridotto. In alcune altre lucertole, l'arco temporale superiore è parzialmente ridotto, mentre nei serpenti (vedi Fig. 86) sono ridotti entrambi gli archi temporali (le ossa frontali posteriori e squamose non si collegano tra loro; entrambi i pozzi temporali rimangono aperti all'esterno).
Pertanto, serpenti e lucertole (squadra squamosa, Squamata) nella struttura del cranio appartengono al gruppo dei rettili diapsidi (a due braccia), ma sono caratterizzati da un diverso grado di riduzione degli archi temporali.

Fig. 85. Cranio di una tartaruga di palude:
1 - una fossa temporale falsa, 2 - un osso mascellare, 3 - un osso mascellare, 4 - un osso zigomatico, 5 - un osso zigomatico quadrato, 6 - un osso quadrato, 7 - un osso squamoso, 8 - un osso frontale, 9 - un osso parietale, 10 - osso frontale, 11 - osso prefrontale, 12 - osso occipitale superiore
La tartaruga ha entrambe le fosse temporali assenti, e la parete laterale del tetto del cranio, delimitando la grande cavità dall'esterno - la cosiddetta fossa temporale falsa (Fig. 85, 1), che si è formata come una rientranza nella parte occipitale del cranio, è composta da ossa densamente fuse: posteriore (post-frontale; Fig. 85, 8), squamoso (squamosum; fig. 85, 7), zigomatico (jugale; fig. 85, 4) e quadrato-zigomatico (quadrato-jugale, fig. 85, 5). Questo tipo di cranio, privo delle vere fosse temporali e degli archi temporali che le limitano, è chiamato anapsidico (inanimato).
Cranio viscerale. Nella lucertola monitor, la cartilagine quadrata palatina si ossifica, formando un osso quadrato nella parte posteriore (quadratum; Fig. 83, 22), all'estremità inferiore a cui è attaccata la mascella inferiore; l'estremità superiore dell'osso quadrato è articolata in modo mobile con un cranio assiale. Di fronte all'osso quadrato si trova il pterigoideo (pterigoideo; Fig. 83, 23), e di fronte c'è l'osso palatino (palatino; Fig. 83, 24), che si collega alle ossa mascellari e al vomere. Tutte queste ossa sono accoppiate; di questi, solo le ossa quadrate hanno origine cartilaginea (primaria).
Dall'osso pterigoideo, il pterigoideo superiore (epipterygoideum; Fig. 83, 25) si estende verso l'alto. Questo osso accoppiato che collega le ossa pterigoideo e parietale è omologa al processo verticale ("ascendente") della cartilagine quadrata palatina ed è caratteristico dei rettili viventi per lucertole e gatteria. Oltre al pterigoideo superiore, le ossa trasversali (trasversale; Fig. 83, 26) si estendono dalle ossa pterigoidi, che nella loro parte anteriore si uniscono alle ossa mascellari. La mascella secondaria è rappresentata dalle ossa mascellari (praemaxillare; Fig. 83, 18) e mascellare (mascellare; Fig. 83, 19). La mascella inferiore è costituita dall'osso articolare primario (articolare; Fig. 83, 30) e dalle ossa tegumentarie: dente (dentale; Fig. 83, 28), angolare (angolare; Fig. 83, 29), sopra-angolare (sopraangulare; Fig. 83, 27), coronarico (coronare; Fig. 83, 31) e, a volte, diverse altre ossa piccole.
Sulle ossa premaxillari, mascellari e dentali dei rettili (ad eccezione delle tartarughe) ci sono semplici denti posteriori conici, a volte leggermente piegati, che crescono fino al bordo dell'osso corrispondente.
L'arco sublinguale, come quello degli anfibi, perse completamente la funzione della sospensione. L'elemento superiore dell'arco ioidico (iomandibolare) fa parte dell'orecchio medio sotto forma di un ossicino uditivo a forma di bastoncino: le staffe (stapes seu columella) e il resto di esso, insieme ai resti degli archi frontali delle branchie, formano l'apparato ioidico.
La struttura descritta del cranio viscerale è generalmente tipica di tutti i rettili. Ma in alcuni gruppi ci sono deviazioni da questo schema, principalmente legate alle specificità della biologia di questi gruppi.

Fig. 86 Cranio di un serpente velenoso:
1 - l'osso mascellare, 2 - l'osso mascellare. 3 - osso palatino, 4 - osso pterigoideo, 5 - osso trasversale, 6 - osso quadrato, 7 - osso squamoso, 8 - osso posteriore, 9 - dente velenoso, 10 - osso frontale, 11 - osso nasale, osso a 12 denti. 13 - osso angolare, 14 - osso articolare
Nei serpenti, non solo le ossa quadrate (fig. 86, 6), ma anche squamose (fig. 86, 7) ad esse collegate, così come le ossa pterigoidi (fig. 86, 4) e palatine (fig. 86, 3) sono molto mobili. Gli ultimi due hanno i denti aguzzi. Le ossa trasversali (Fig. 86, 5) nei serpenti servono come leve, per così dire, trasmettendo il movimento delle ossa pterigoidi alle ossa mascellari (Fig. 86, 2), che a loro volta sono molto mobili. Questo intero sistema di ossa articolate in modo mobile non solo contribuisce a un'apertura estremamente ampia della bocca, ma fornisce anche movimenti indipendenti delle metà destra e sinistra dell'apparato mascellare quando si spinge la preda nella gola con intercettazione alternata. Ciò consente ai serpenti di ingoiare una preda relativamente molto grande (che supera lo spessore del corpo del serpente). I serpenti velenosi sulle ossa mascellari hanno denti velenosi affilati mobili, curvi all'indietro (Fig. 86, 9) che hanno un canale interno o una scanalatura sulla superficie anteriore, attraverso la quale il veleno scorre dalle ghiandole velenose situate alla base del dente quando viene morso.
Il teschio di coccodrillo (Fig. 84) è caratterizzato dal fatto che i denti non crescono fino al bordo delle ossa dentali, mascellari e mascellari, come in altri rettili, ma siedono in cavità speciali (fori o alveoli) di queste ossa - denti tecodontici. Un'altra caratteristica del cranio viscerale dei coccodrilli è un palato duro secondario che separa la cavità orale dal passaggio rinofaringeo.
I processi palatino delle ossa premaxillari (Fig. 84,7) e mascellare (Fig. 84, 2), così come le ossa palatina (Fig. 84, 17) e pterigoideo (Fig. 84, 18), prendono parte alla formazione del palato duro secondario. A causa della formazione di un palato duro, i cigni secondari (Fig. 84, 20) vengono portati indietro e situati nelle ossa pterigoidi, sopra la laringe. La formazione di un palato duro secondario è collegata alla natura dello stile di vita del coccodrillo: il contatto diretto della laringe con il coro apre la possibilità di respirazione ininterrotta quando si mangia e quando il coccodrillo riposa in acque poco profonde, esponendo le narici sollevate dall'acqua, mentre la cavità orale è piena d'acqua.
Arti accoppiati e loro cinture. Cinturone i rettili sono costituiti da ossa tipiche: la scapola situata più dorsalmente (scapola; Fig. 87, 1) e la coracoide (coracoideum; Fig. 87, 3) rivolta verso il lato ventrale. Entrambe queste ossa sono coinvolte nella formazione della fossa articolare (Fig. 87, 4) per attaccare la zampa anteriore. Dorsale alla scapola è una cartilagine soprascapolare spianata (cartilago suprascapularis; Fig. 87, 2), e davanti alla corteccia è procoracoide cartilagineo (cartilago procoracoidea; Fig. 87, 5). C'è uno sterno ben sviluppato (sterno, Fig. 87, 6), a cui sono attaccate diverse costole (costa, Fig. 87, 7). Pertanto, a differenza degli anfibi, il rettile sviluppa un torace e la cintura della spalla ha supporto nello scheletro assiale. Sul lato ventrale dello sterno vi è un osso tegumentario a forma di T - suprasternum (episterno; Fig. 87, 8), di fronte ad esso - anche ossa tegumentarie - clavicola (clavicola; Fig. 87, 9). Le estremità esterne della clavicola sono attaccate alle scapole e l'interno fuso con i rami del torace. Le clavicole e il sovrasternum (assenti negli anfibi) aumentano la forza di legame delle parti destra e sinistra della cintura della spalla.

Fig. 87. La cintura della spalla della lucertola monitor (vista dal basso):
1 - scapola, 2 - cartilagine soprascapolare, 3 - coracoide, 4 - cavità articolare per la testa della spalla, 5 - cartilagine pro-toracica, 6 - sterno, 7 - costole, 8 - sovrasternale, 9 - clavicola
Nei serpenti, la cintura della spalla è completamente ridotta, e nelle tartarughe, le clavicole e il sovrasternum sono inclusi nelle ossa dello scudo addominale (plastron; vedi Fig. 82, B) del carapace, formando rispettivamente le placche ossee accoppiate anteriori e non accoppiate tra loro.

Fig. 88. Cintura pelvica di lucertola monitor (vista dal basso):
1 - ilio, 2 - osso pubico, 3 - osso sciatico, 4 - acetabolo (fossa articolare) per la testa del femore, 5 - vertebre sacrali
La cintura pelvica è costituita da due metà simmetriche collegate lungo la linea mediana della cartilagine. Ogni metà è composta da tre ossa; situato dorsalmente iliaco (ilio; Fig. 88, 1), situato sul lato ventrale del pube (pube; Fig. 88, 2) e sciatico (ischio, Fig. 88, 3). Tutte queste ossa prendono parte alla formazione della fossa articolare (acetabolo; Fig. 88, 4), a cui è attaccato l'arto posteriore. Il bacino del rettile è chiuso: le ossa pubica e ischiale destra e sinistra sul lato ventrale sono fuse insieme.

Fig. 89. Arti della lucertola monitor
A - fronte; B - indietro:
1 - omero, 2 - ulna, 3 - raggio, 4 - polso, 5 - metacarpo, 6 - falange delle dita, 7 - articolazione intercarpale, 8 - femore, 9 - tibia, 10 - tibia, 11 - patella, 12 - tarso, 13 - articolazione intertarsale, 14 - metatarso
Gli arti dei rettili sono costruiti secondo il modello tipico degli arti dei vertebrati terrestri. La parte prossimale dell'avambraccio (Fig. 89, A) è rappresentata da un osso - l'omero (omero; Fig. 89, 1), seguito dall'avambraccio, che consiste di due ossa - l'ulna (ulna; Fig. 89, 2) e il raggio (raggio; Fig. 89, 3). Il polso (carpo, Fig. 89, 4) è costituito da ossa relativamente piccole, solitamente situate in due file; sul lato di loro è un altro osso a forma di pera, preso per il resto del sesto dito. Il metacarpo (metacarpo; Fig. 89, 5) è composto da cinque ossa allungate a cui sono attaccate le falangi delle cinque dita (falange digitorum; Fig. 89, 6).
Le ultime falangi portano artigli. L'articolazione, fornendo mobilità della mano, nei rettili non passa tra le ossa dell'avambraccio e la fila prossimale delle ossa del polso (come negli anfibi), ma tra le file prossimale e distale delle ossa del polso. Tale articolazione è chiamata intercarpale (Fig. 89, 7).
Nell'arto posteriore, l'elemento prossimale - il femore (femore; Fig. 89, 8) è articolato articolazione del ginocchio con una tibia composta da due ossa di tibia: grande (tibia; Fig. 89, 9) e piccola (fibula; Fig. 89, 10). Sopra la superficie anteriore di questa articolazione è presente un piccolo osso: la rotula (rotula; Fig. 89, 11). Nel tarso (tarso; Fig. 89, 12), la fila prossimale di ossicini si fonde o si collega quasi immobile alle ossa della parte inferiore della gamba, e anche gli ossicini della fila distale sono strettamente collegati e parzialmente fusi con le ossa del metatarso. Per questo motivo, la superficie articolare qui si trova non tra la parte inferiore della gamba e del piede, ma tra le file prossimale e distale delle ossa tarsali. Tale articolazione è caratteristica dei rettili ed è chiamata articolazione intertarsale (Fig. 89, 13). Il metatarso (metatarso; Fig. 89, 14) è costituito da cinque ossa allungate a cui sono attaccate le falangi delle cinque dita (falangi digitorum; Fig. 89, 6). Le falangi terminali portano artigli.
CONCLUSIONE
Rettili (Reptilia) - la prima classe del gruppo di veri vertebrati terrestri (Amniota), i cui rappresentanti hanno un complesso di adattamenti per vivere sulla terra senza alcun legame con l'ambiente acquatico.
Si è detto sopra che la stretta connessione degli antenati dei rettili - anfibi - con corpi idrici (o almeno con luoghi di elevata umidità) era dovuta principalmente a due caratteristiche biologiche: il tipo di riproduzione acquatica e la dualità del sistema respiratorio (polmoni e pelle). Queste caratteristiche si riflettono a loro volta sulla natura del metabolismo dell'acqua: vivendo in un ambiente umido, gli anfibi non hanno meccanismi morfofisiologici per ridurre la perdita di umidità del corpo. Il pieno sviluppo della terra è impensabile senza trasformazioni fondamentali come la riproduzione, la respirazione e il metabolismo dell'acqua.
L'implementazione di queste trasformazioni caratterizza la classe dei rettili.
Le caratteristiche più specifiche di questa classe (e di altri amnioti) come vertebrati terrestri primari sono associate all'adattamento alla vita nell'aria atmosferica relativamente secca. Le caratteristiche strutturali specifiche dei rettili sono principalmente associate all'adattamento a un uso più economico dell'umidità. La pelle dei rettili è secca e priva di ghiandole (ad eccezione di un piccolo numero di ghiandole specializzate che secernono un segreto relativamente spesso). Gli strati esterni dell'epidermide sono rappresentati da cellule morte piene di sostanza cornea (cheratogialina).
Questo strato corneo protegge il corpo dall'essiccamento e migliora anche notevolmente la protezione da danni chimici e meccanici. Sotto lo strato corneo, lo strato malpigianale dell'epidermide è rappresentato da cellule viventi. Man mano che le cellule morte si consumano, vengono sostituite da cellule degli strati inferiori, nel protoplasma di cui compaiono inizialmente dei granuli di cheratogialina separati, che successivamente riempiono l'intera cellula. Oltre alla cheratinizzazione generale dello strato superiore dell'epidermide, i rettili sono caratterizzati dalla formazione nella pelle di varie forme di squame cornee che coprono la superficie della pelle. Le squame forniscono protezione meccanica della pelle e contribuiscono a una riduzione ancora maggiore della perdita di umidità attraverso il tegumento; alcuni rettili moderni (ad esempio i coccodrilli) hanno placche ossee sotto le squame cornee.
Tutte queste trasformazioni, biologicamente benefiche in termini di mantenimento di un bilancio idrico più economico del corpo, sono state possibili principalmente perché la pelle dei rettili non partecipa alla respirazione, e ciò è dovuto al progressivo sviluppo del sistema respiratorio (vedi sotto).
Di grande importanza nell'evoluzione dei vertebrati terrestri superiori è la trasformazione del sistema escretore. Contrariamente alle classi precedentemente studiate, i rettili (come altri Amniota) possiedono reni pelvici (metanephros). A differenza dei reni del tronco (mesonefrosi), l'anamnesi dei tubuli escretori del rene pelvico ha perso completamente il contatto con la cavità corporea; i prodotti metabolici entrano nei tubuli escretori solo dal plesso dei capillari sanguigni - glomerulo Malpighiano. È racchiuso in una sporgenza vuota dell'estremità del tubulo - la capsula di Bowman. Ciò porta al fatto che nel processo di isolamento dei prodotti di dissimilazione, il corpo perde meno acqua (il fluido della cavità non viene escreto). I vasi sanguigni che emergono dal glomerulo malpighiano formano una seconda rete di capillari attorno alla parete del tubulo escretore, e poi scorrono nei vasi più grandi del sistema venoso. Attraverso questa seconda rete di capillari, acqua, carboidrati e alcuni sali vengono riassorbiti nel sangue, in modo che la cosiddetta "urina secondaria" che viene rilasciata diventa più concentrata e ha una composizione diversa rispetto all '"urina primaria" formata nelle capsule di bowman.
Il rene pelvico (metanephric) ha il suo uretere, non omologa ai canali Müller o Wolf. Il canale del lupo è conservato solo nei maschi, dove è collegato attraverso l'epididimo alla ghiandola genitale e svolge il ruolo di vas deferens. Il canale di Mueller nei maschi è ridotto. Le femmine, d'altra parte, mantengono solo i canali di Müller; svolgono la funzione degli ovidotti e si aprono nella cavità del corpo con fori - imbuti degli ovidotti. I canali del lupo nelle femmine sono ridotti.
Pertanto, le caratteristiche principali del sistema escretore dei vertebrati terrestri superiori sono la perdita di comunicazione tra i reni e la cavità corporea e il rafforzamento del sistema di assorbimento inverso dell'acqua dall'urina primaria nel sangue. Queste proprietà garantiscono l'implementazione della funzione escretoria con un consumo minimo di acqua, che per gli abitanti della terra, come già accennato, ha un grande significato biologico. A ciò possiamo aggiungere che tra i vertebrati terrestri, l'intensità della filtrazione generale e l'assorbimento inverso dell'acqua è diversa negli animali che vivono in condizioni diverse. Quindi, tra i rettili, i coccodrilli e le tartarughe (principalmente forme d'acqua dolce) hanno glomeruli ben sviluppati e secernono urina liquida, mentre in lucertole e serpenti, i glomeruli sono notevolmente più deboli e l'urina è solida o semi-liquida.
L'intero complesso delle caratteristiche considerate, fornendo un bilancio idrico più economico dei rettili, è alla base della separazione finale dei vertebrati terrestri superiori dall'ambiente acquatico e della loro popolazione diffusa di vari biotopi terrestri.
Insieme a queste caratteristiche, i rettili sono caratterizzati da una serie di trasformazioni che hanno un valore progressivo comune nell'evoluzione dei vertebrati.
Il sistema respiratorio è cambiato in modo significativo. Apparve una trachea, la cui lunghezza dipende dalla lunghezza del collo. I polmoni dei rettili sono caratterizzati da una struttura interna più complessa rispetto agli anfibi: hanno una traversa interna che li divide in dipartimenti. Da queste traverse, a loro volta, partono le partizioni, sviluppate a diversi gradi in diversi gruppi. Tra i setti si formano le vie respiratorie intrapolmonari, che fungono da continuazione dei bronchi, e in alcuni casi sono divise in bronchi intrapolmonari del secondo ordine. Le pareti interne delle camere formate da questo intero sistema di travi hanno una struttura cellulare, in modo che di conseguenza i polmoni acquisiscano generalmente una struttura spugnosa complessa. Un tale dispositivo dei polmoni aumenta significativamente la superficie respiratoria totale e, di conseguenza, l'intensità della respirazione. Grazie a questi dispositivi, il fabbisogno di ossigeno dell'organismo dei rettili è completamente fornito dalla sola respirazione polmonare.
È estremamente importante che l'atto di respirare i rettili sia realizzato espandendo e contraendo attivamente la cavità addominale sotto l'azione dei muscoli addominali e costali. Questo tipo di respirazione fornisce una ventilazione più intensiva dei polmoni con un grande volume d'aria; il suo aspetto è associato allo sviluppo di un torace mobile.
Trasformazioni progressive significative si verificano nel sistema circolatorio dei rettili (nella descrizione del sistema circolatorio è stato fornito uno schema per la separazione dei flussi di sangue venoso e arterioso nel cuore dei rettili).
A causa della comparsa di un setto incompleto nel ventricolo e del passaggio indipendente da diversi reparti dei principali tronchi arteriosi (che è associato a una riduzione del cono arterioso), si ottiene una separazione abbastanza completa dei flussi sanguigni arterioso e venoso. Al momento della sistole ventricolare, le sue cavità interne sono isolate per qualche tempo; mentre il sangue venoso entra nell'arteria polmonare, il sangue arterioso entra nell'arco aortico destro e la miscela venosa arteriosa entra nell'arco aortico sinistro. Poiché entrambe le arterie carotide e succlavia provengono dall'arco aortico destro, la testa e la parte anteriore del corpo sono alimentate con sangue arterioso quasi puro (ad esempio, nelle tartarughe nelle arterie carotidi la saturazione di ossigeno nel sangue ha superato il 90% nella metà dei casi). I vasi che si estendono sotto la connessione degli archi aortici trasportano il sangue contenente una certa impurità venosa agli organi della metà posteriore del corpo.
Il setto orizzontale che divide il ventricolo del cuore del rettile in due camere principali non è omologa al setto interventricolare di uccelli e mammiferi. Di tutti i gruppi di rettili solo nei coccodrilli, un nuovo setto - verticale - appare nel ventricolo, sostituendo l'antico orizzontale. Questo setto verticale divide completamente il ventricolo cardiaco dei coccodrilli in due camere isolate: la destra (venosa) e la sinistra (arteriosa). Pertanto, il cuore dei coccodrilli può essere chiamato a quattro camere: ha due atri isolati l'uno dall'altro e due ventricoli anche completamente separati. Tuttavia, insieme alla completa separazione dei ventricoli, i coccodrilli mantengono entrambi gli archi aortici, che formano una croce caratteristica di tutti i rettili: la destra inizia a sinistra e la sinistra nel ventricolo destro. All'incrocio c'è una cosiddetta apertura panoramica che collega le cavità di entrambi gli archi aortici.
Le arterie carotide e succlavia, come tutti i rettili, si estendono dall'arco aortico destro. L'arco aortico sinistro, che circonda il cuore, dà vasi sanguigni all'intestino e, attraverso un condotto sottile, si collega all'arco aortico destro, dando origine all'arteria spinale. Secondo alcuni rapporti, alla base dell'arco aortico sinistro ci sono valvole e un sistema di cartilagine che impedisce l'ingresso di sangue venoso con la sistole ventricolare.
Alcuni autori spiegano la conservazione di entrambi gli archi aortici e la loro connessione nel sito di intersezione con le caratteristiche ecologiche dei coccodrilli: mentre sott'acqua, la circolazione polmonare fa circolare un po 'di sangue, perché la respirazione è sospesa e il sangue in eccesso dalla circolazione polmonare passa in un grande cerchio, sostenendo così i suoi vasi sanguigni pressione sanguigna normale. È possibile che anche la conservazione dei dotti botallali (vasi che collegano gli archi aortici con le arterie polmonari) nelle tartarughe che conducono uno stile di vita acquatico sia della stessa importanza. Come i dotti carotidi (la connessione degli archi aortici con le arterie carotidi) di serpenti e lucertole, queste formazioni sono i rudimenti degli archi arteriosi dei lontani rettili ancestrali, che sono stati ridotti da un cambiamento nella respirazione delle branchie dei polmoni. Il più primitivo dei rettili viventi, l'hatteria, ha dotti sia carotidi che duttali.
centrale sistema nervoso i rettili sono caratterizzati da un aumento delle dimensioni relative del cervello e dal progressivo sviluppo degli emisferi del cervello anteriore, nel cui tetto appare la materia grigia superficiale - appare per prima la corteccia cerebrale (ancora debolmente sviluppata).
Tra le caratteristiche progressive dello scheletro dei rettili, l'aspetto di un torace chiuso è importante. Oltre al valore già indicato di questa trasformazione nell'intensificazione della respirazione, il torace assicura la connessione della cintura della spalla con lo scheletro assiale, e quindi la sua fissazione più solida, che porta ad un aumento funzione di supporto arti. Nei serpenti che hanno perso gli arti accoppiati per la seconda volta, anche le cinture sono ridotte.
Grazie all'unico condilo occipitale, la struttura specifica delle prime vertebre cervicali, che consente agli atlanti di ruotare attorno al processo simile a un dente dell'epistrofia, e la comparsa di molte vertebre cervicali che sono articolate in modo mobile l'una con l'altra, la mobilità della testa del rettile è notevolmente aumentata. Questo è di grande importanza biologica nell'orientamento, nella ricerca e nell'ottenimento di cibo, ecc.
I cambiamenti evolutivi nel tetto del cranio sono molto significativi. Nei rettili più antichi, le pareti superiore e laterale del cranio erano composte da numerose ossa tegumentarie e avevano l'aspetto di un solido carapace osseo. Un tale teschio era chiamato trapuntato. Un'ulteriore evoluzione ha seguito il percorso di graduale riduzione delle ossa tegumentarie del cranio trapuntato in modo tale che si formassero depressioni (fosse temporali) al posto delle ossa ridotte, delimitate da ponti ossei relativamente sottili (archi temporali). Queste trasformazioni del cranio, da un lato, hanno portato al suo sollievo, che è importante quando si vive in un ambiente molto meno denso dell'aria acquatica; d'altra parte, venivano usate depressioni nel sito delle ossa ridotte del tetto del cranio. Per accogliere i muscoli della mascella. Lo sviluppo dei muscoli della mascella è direttamente correlato ad un aumento dell'attività nutrizionale.
Nella maggior parte dei gruppi di rettili durante la riduzione del tetto secondario del cranio, si sono formate due fossa temporali: quella superiore, delimitata esternamente dall'arco temporale superiore (ossa frontali e squamose posteriori), e quella laterale, delimitata sopra l'arcata temporale superiore e esterna (ossa zigomatiche e zigomatiche) inferiori. I rettili con questo tipo di cranio appartengono al gruppo del diapside (bipede). In alcuni rappresentanti di questo gruppo, la riduzione ossea è andata ulteriormente nella direzione della perdita di uno o di entrambi gli archi temporali. Un cranio diapside tipicamente pronunciato è caratteristico delle forme viventi di hatteria e coccodrilli. In molte lucertole, l'arco temporale inferiore è parzialmente ridotto e in alcuni quello superiore. Entrambi gli archi temporali sono ridotti in serpenti. Gli uccelli si sono evoluti da rettili diapsidi (in questi animali l'arco temporale inferiore è stato completamente preservato, ma quello superiore è stato ridotto).
In alcuni rettili, l'evoluzione della regione temporale del cranio ha seguito il percorso di formazione di una sola fossa temporale, limitato a un singolo arco temporale composto da ossa zigomatiche e squamose. Poiché l'osso zigomatico è un elemento caratteristico della parte inferiore e l'osso squamoso è l'arco temporale superiore del cranio diapside, un tale arco viene chiamato misto e i rettili con tale struttura cranica appartengono al gruppo sinapsid (rettili con un arco misto). Tra i rettili moderni, non ci sono forme sinapsidali. Questo tipo di cranio è caratteristico dei discendenti dei rettili sinapsidi: i mammiferi.
In uno dei rami dei rettili primitivi, la riduzione delle ossa tegumentarie del tetto del cranio è andata diversamente: la formazione di una tacca nella parte occipitale del cranio tegumentario ha portato alla formazione su ciascun lato di una cavità - una falsa fossa temporale, coperta all'esterno con un ampio ponte osseo dall'osso anteriore, squamoso e zigomatico quadrato. Tali rettili appartengono al gruppo di anapsidi (irragionevoli). Dei rettili viventi, le tartarughe appartengono a questo gruppo.
L'insieme delle caratteristiche progressive dei rettili ha portato ad un aumento generale dell'energia vitale dell'attività di questi animali e alla comparsa tra loro di un'ampia varietà di forme di vita, che riflette gli adattamenti alle varie condizioni di vita. A differenza degli anfibi, che mangiano esclusivamente cibo per animali, tra i rettili ci sono forme erbivore (come, ad esempio, le tartarughe terrestri). Tuttavia, i rettili più moderni sono carnivori. Una maggiore mobilità rispetto agli anfibi consente loro di cacciare prede mobili, raggiungendo talvolta dimensioni significative. In diversi casi, il successo della caccia è facilitato da una serie di dispositivi speciali: denti velenosi in alcune specie di serpenti, il colore protettivo che caratterizza molti rettili, ecc.
La maggior parte dei rettili sono animali terrestri. Sulla terra, diversi tipi di rettili si trovano in una grande varietà di condizioni: nei deserti, boschetti di vegetazione palustre, foreste, tra rocce, ecc. Tutte queste specie hanno dispositivi adeguati per il movimento (dita delimitate da piastre di corno in deserti di sabbia o setole di corno che consentono forme di arrampicata rimanere su superfici ripide) per evitare gli attacchi dei predatori (scavare nella sabbia, autotomia della coda, ecc.), ecc.
Molti rettili sono nuovamente passati alla vita acquatica (tartarughe marine e d'acqua dolce, coccodrilli, serpenti acquatici). Alcune di queste specie trascorrono quasi tutta la loro vita in acqua, ma vanno necessariamente a terra per la riproduzione. I serpenti marini hanno sviluppato la capacità di ovipare, il che ha permesso loro di tornare completamente allo stile di vita acquatico (tra i rettili estinti, gli ittiosauri, anch'essi dotati della capacità di ovipare), ha condotto uno stile di vita completamente acquatico).
I rettili raggiunsero la più grande diversità di forme di vita durante il periodo di massimo splendore evolutivo di questa classe - nell'era mesozoica. Tra rettili fossili, erbivori e carnivori (diversi gruppi di dinosauri), sono noti abitanti profondamente specializzati dell'ambiente acquatico (ittiosauri) e forme volanti (pterodattili, ramphorinch), così come molti altri gruppi biologici. Il raffreddamento generale del clima che è seguito al Mesozoico non ha contribuito all'ulteriore prosperità dei rettili: come gli anfibi, i rettili sono poikilotermici e non possono mantenere a lungo un'alta temperatura corporea durante lo schiocco freddo.
La maggior parte delle specie di rettili moderni vive in climi caldi; le specie comuni nella zona temperata trascorrono una parte significativa dell'anno in stato di stordimento. In queste condizioni, chiari vantaggi si sono rivelati dalla parte dei loro "discendenti" - uccelli e mammiferi, nella cui evoluzione è apparsa una nuova preziosa qualità - l'omeoterma. La capacità di mantenere la vita attiva in una vasta gamma di temperature ambientali è uno dei motivi principali per determinare la posizione prevalente di uccelli e mammiferi nella moderna fauna dei vertebrati.
Ulteriori letture
Gurtovoi N.N., Matveev B.S. Dzerzhinsky F. Ya. Pratica zootomia dei vertebrati. Anfibi, rettili. M., 1978
Terentyev P.V. Herpetology. M., 1961.
Schmalhausen I. I. Fondamenti dell'anatomia comparata dei vertebrati. M., 1947
La colonna vertebrale combina 4 dipartimenti: cervicale, lombare-toracico, sacrale e caudale. I corpi vertebrali sono concavi nella parte anteriore e convessi nella parte posteriore - intere vertebre.
Gli archi superiori terminano con un processo spinoso ben definito; una coppia di processi articolari anteriori corti parte dal margine anteriore dell'arco superiore e una coppia di processi articolari posteriori parte dal bordo posteriore dello stesso arco. Non ci sono segni di residui di accordi negli adulti.
Scheletro di lucertola (secondo Bram):
1 - clavicola, 2 - scapola, 3 - spalla, 4 - raggio, 5 - ulna, 6 - polso, 7 - polso, 8 - falange delle dita, 9 - costole, 10 - bacino, 11 - coscia, 12 - grande tibia, 13 - tibia, 14 - tarso, 15 - metatarso, 16 - vertebre caudali
rachide cervicale rappresentato da otto vertebre. La parte anteriore di essi, chiamata atlante, o atlante, ha una forma ad anello caratteristica di tutti gli amnioti. Sul lato anteriore della sua parte inferiore c'è una superficie articolare spaiata, con l'aiuto della quale la vertebra si articola con il cranio. Il foro dell'atlante è diviso da un fascio in due metà: superiore e inferiore. Il midollo spinale passa attraverso il foro superiore e il processo dentoide (processus odontoideus) entra nel foro inferiore, che è saldamente collegato, ma non fuso alla seconda vertebra, chiamata epistropheus. Come dimostrano i dati embriologici, il dentato è il corpo distaccato della prima vertebra. Pertanto, l'atlante è mobile rispetto al proprio corpo. Le ultime tre vertebre cervicali portano una coppia di costole cervicali ben sviluppate, che sono attaccate in modo mobile ai corpi vertebrali, aumentano gradualmente posteriormente e terminano liberamente senza raggiungere lo sterno. In generale, la parte cervicale dei rettili differisce da quella degli anfibi non solo in un gran numero di vertebre, ma anche in una più mobile: articolazione della prima vertebra sia con il cranio che con la seconda vertebra. Di conseguenza, la testa riceve molta più libertà di movimento.
colonna lombare-toracica la lucertola contiene 22 vertebre. Tutti sono dotati di nervature mobili, la cui lunghezza diminuisce gradualmente verso la parte posteriore. Ogni costola è costituita dalla parte superiore ossea e dalla cartilagine inferiore. Le prime cinque vertebre lombari-toraciche (in realtà le vertebre toraciche) hanno costole particolarmente lunghe che si attaccano allo sterno, formando torace (Torace). Lo sterno (sterno) è cartilagineo e ha la forma di una placca rombica. Lo sviluppo embrionale dello sterno di amniot mostra che è formato dall'intercrescita del distaleestremità delle costole e, pertanto, non è omologa allo "sterno" degli anfibi, formato indipendentemente. Pertanto, nella lucertola, è già delineata una divisione del tronco della colonna vertebrale nella colonna toracica e lombare, che è estremamente caratteristica degli amnioti.
Sezione di coda è costituito da numerose vertebre (diverse decine). Quelle anteriori sono simili a quelle sacrali, ma i loro corpi sono più lunghi, i processi trasversali sono più sottili e i processi spinosi sono più lunghi. Verso la parte posteriore, le vertebre caudali diminuiscono gradualmente di dimensioni, perdono i loro processi e si trasformano in piccole ossa a forma di bastoncino.
Articoli più interessanti
Lo sviluppo e la complicazione del sistema motorio dei rettili è causato dalla transizione verso un habitat predominante sulla terra e dalla necessità di superare le irregolarità del suo rilievo. I principali cambiamenti nell'apparato di supporto dei rettili, rispetto agli anfibi, sono associati maggiore mobilitànecessario per la ricerca riuscita di cibo, riproduzione, protezione dai pericoli, ecc. Ciò si riflette in indurimento del cranio, maggiore differenziazione dello scheletro assiale, rafforzamento degli arti e delle loro cinture, ulteriore sviluppo di muscoli specializzati.
Lo scheletro dei rettili è caratterizzato da sviluppo significativo dell'osso elementi e correlati riduzione della cartilagine formazioni. Un numero particolarmente elevato di ossificazioni tegumentarie compone il tetto, i lati e il fondo del cranio cerebrale. Cambiamenti significativi e reparto viscerale. L'apparato mascellare è diventato più massiccio, le ossa della mascella superiore sono profondamente radicate parte superiore dipartimento del cervello. Allo stesso tempo, il cranio ha acquisito mobilità nella connessione delle sezioni cerebrali e viscerali. Quindi, i serpenti fondo il cranio è un complesso sistema a leva articolata, che ti consente di ingoiare prede intere di grandi dimensioni (boa, serpenti a uovo, ecc.). Contribuisci a questo fosse e archi temporali, essere un luogo di attaccamento di un forte e muscoli elastici. Lo scheletro della testa si articola con la colonna vertebrale attraverso condilo singoloformato dalle ossa della nuca. In generale, il cranio è diventato più forte e, allo stesso tempo, più leggero.
La complicazione della differenziazione della colonna vertebrale si manifesta in un aumento del numero di vertebre nei dipartimenti del tronco. Di particolare importanza allargamento cervicale dipartimento, che determina la mobilità della testa, che, a sua volta, migliora l'orientamento a terra e aiuta ad accelerare il movimento. Fondendo la parte anteriore delle costole con lo sterno, si forma torace. Si sviluppa nella zona del torace muscoli intercostali, che determina il meccanismo respiratorio in molte specie.
estremità la maggior parte dei rettili sono corti, attaccati alle cinture degli arti e hanno muscoli specializzati. Nell'area di connessione dell'avambraccio con la spazzola si forma articolazione intercarpale. Una formazione simile ha luogo negli arti posteriori, dove si forma la parte della caviglia articolazione intertarsale. Queste formazioni contribuiscono in modo significativo al rafforzamento e alla mobilità degli arti inferiori. Cintura degli arti pur mantenendo una notevole quantità di cartilagine, gli arti sono attaccati a scheletro assiale.
Rettili che hanno perso arti, cinture e petto (serpenti, lucertole senza gambe), hanno costole lunghe, tronco forte e muscoli sottocutanei. In un numero di rettili (tartarughe), la perdita di velocità di movimento è associata allo sviluppo di uno speciale dispositivo di protezione - guscio fissomovimento a catena. Sono presenti anche le caratteristiche strutturali dello scheletro specie acquatiche secondarie (tartarughe marine, serpenti marini, iguane, coccodrilli).
Dopo aver familiarizzato con le caratteristiche di base dello scheletro, possiamo procedere a un esame più dettagliato della struttura delle sue parti costituenti.
cranio
Il cranio è allungato, con un contenuto significativo di elementi ossei (Fig. 51). Viene determinata una forma particolare del cranio, il rapporto tra le dimensioni delle sue parti costituenti in diversi rettili tipo di alimentazione e metodo di acquisizione cibo.
Cranio del cervello indurito aumentando il numero di ossa e la loro solida fusione tra loro.
Il tetto teschi formati da accoppiati ossa nasali, frontali anteriori, frontali, posteriori (postorbitale) e parietali. impareggiabile osso interparietal ha forame parietale autorità. Ai lati di questi elementi crescono ossa lacrimali e infraorbitaliche incornicia la parte superiore dell'orbita degli occhi.
fondo forma di teschi osso sfenoide principale con rudimentale parasphenoid e apriscatoleai cui lati si trovano coane. Gli elementi della sezione viscerale prendono anche parte alla formazione del fondo del cranio cerebrale. Ossa pterigoidi articolare con piazza, che nei serpenti e nelle lucertole sono collegati in modo mobile al cranio del cervello e fungono da sospensione dell'apparato mascellare. tra pterigoidi e mascellaresi trova osso trasversale, che rafforza ulteriormente il cranio.
occipitale consiste di quattro ossa - principale, sopraoccipitale e laterale accoppiato. Come risultato della loro interscambio, un condilo occipitale. Nella parte inferiore del dipartimento c'è forame occipitale per collegare il cervello e il midollo spinale.
Parte laterale i teschi formano tre paia ossa auricolari (anteroposteriore, auricolare superiore e auricolare posteriore), che sono coinvolti nell'istruzione fosse temporali.Le ossa superiori e posteriori sono fuse con le ossa della regione occipitale. Accanto a loro ci sono temporale, scalaparti superiori alar ossa. accoppiato ossa colonnari collega il tetto del cranio al fondo. Grandi orbite formano elementi ossei adiacenti - infraorbitale, postorbitale, zigomatico, lacrimale. Setto interorbitale formata da escrescenze delle ossa frontali e parietali. Nella parte anteriore del cranio cerebrale le ossa mascellari e intermaxillari si incastrano in alto. olfattivoarea del cranio cartilagine.
Fig. 51. Il cranio di una lucertola sopra (A), sotto (B), lato (C) e mascella inferiore:
1 - il forame occipitale, 2 - l'osso occipitale laterale, 3 - l'osso occipitale superiore, 4 - l'osso occipitale principale, 5 - il condilo occipitale, 6 - l'osso sfenoide principale, 7 - il vomero, 8 - ciano, 9 - il parafenoide vestigiale, 10 - parietale osso, 11 - un'apertura per l'occhio parietale, 12 - l'osso frontale, 13 - l'osso nasale, 14 - l'osso mascellare, 15 - l'osso mascellare, 16 - l'osso prefrontale, 17 - l'osso lacrimale, 18 - l'osso infraorbitale, 19 - l'osso orbitale, 20 - osso squamoso, 21 - osso temporale, 22 - osso zigomatico, 23 - narice, 24 - osso quadrato, 25 - osso palatino, 26 - osso pterigoideo, 27 - osso colonnare, 28 - osso trasversale, 29 - osso articolare, osso a 30 denti, 31 - osso angolare, 32 - l'osso arcuato, 33 - processo coronoideo
Il cranio di un numero di rettili è caratterizzato dalla presenza di ben definiti archi temporali (sono ridotti nella lucertola). Solo alla lucertola monitor archi temporali superiorialla cui educazione prendono parte orbitale e squamoso ossa. Delimitano questi archi laterale fosse temporali. I coccodrilli, insieme agli archi superiori, hanno inferioreche si formano mascellare e zigomaticoossatura (cranio diapsid) (Fig. 52). in tartarughenon ci sono fosse temporali e archi temporali ( cranio di tipo anapside).
Cranio visceraleè composto da mascella e apparato ipoide . ossaturala parte superiore mascella (intermaxillary e mascellare), come già notato sopra, sono stati rafforzati e profondamente radicati nel cranio cerebrale. Ha luogo autostiliya - connessione diretta del cranio viscerale con il cervello. Come risultato dell'ossificazione e della trasformazione della cartilagine quadrata palatina quadrato e colonnare ossa. Palatino e pterigoideo gli elementi sono di origine cutanea e sono coinvolti nella formazione del fondo del cranio cerebrale.
Numerosi rettili (coccodrilli, tartarughe) hanno un posto palato osseo secondario, che è stato formato dalla crescita di un certo numero di ossa - intermaxillary, mascellare, palatino e alar. I coccodrilli sono cambiati e posizione della narice: le narici esterne sono elevate sopra la superficie della testa e le choana vengono spostate nella regione occipitale del cranio. Queste trasformazioni sono alla base della separazione del rinofaringe dalla cavità orale, che consente agli animali di tenere la bocca aperta sott'acqua e allo stesso tempo respirare attraverso le narici esposte alla superficie dell'acqua.
Mascella inferiore formata da ossificazione cartilagine di Meckel e apparenze articolato, dentale, angolare, arcuato, coronale ossa e un numero di piccole formazioni. Delle ossa elencate, solo l'articolare ha un'origine condrale, tutto il resto è tegumentario.
Apparato sublinguale rappresenta record (copula omologa) con tre paia di corna - derivati \u200b\u200bdi hyoids e archi ramificati. Iomandibolare, come negli anfibi, sotto forma di ossicino uditivo ( staffa)situato nell'orecchio medio.
Attrezzature e materiali
1. Scheletri di una lucertola, tartarughe, serpenti (un set per due studenti).
2. Vertebre di lucertola provenienti da diverse parti del corpo.
3. Gli arti anteriori e posteriori della lucertola con cinture.
4. Cranio di una lucertola, tartarughe, serpenti.
5. Tabelle: 1) lo scheletro di una lucertola; 2) la struttura delle vertebre da diverse parti del corpo; 3) cranio (vista dall'alto e dal basso); 4) lo scheletro degli arti e le loro cinture.
Osservazioni introduttive
La direzione principale dell'evoluzione dei rettili è stata quella di migliorare l'apparato locomotore e aumentare la diversità dei movimenti. I rettili (moderni ed estinti) dimostrano tutti i principali modi di movimento noti ai vertebrati.
Il modo di movimento evolutivamente originale e tipico è stato preservato nelle lucertole moderne. Come per gli anfibi muniti di coda, il movimento viene effettuato alternativamente riorganizzando gli arti relativamente corti posti ai lati
quando si piega il corpo a destra - a sinistra. La struttura delle articolazioni degli arti e il modello generale di locomozione nelle lucertole moderne sono solo leggermente modificati rispetto ai primi rettili.
L'efficacia di questo metodo di movimento tra i rettili è diventata più elevata rispetto agli anfibi. Ciò è stato ottenuto, da un lato, rafforzando e differenziando i muscoli del corpo e degli arti, e dall'altro, differenziando lo scheletro e rafforzando e trasformando i suoi singoli nodi e articolazioni. Rafforzare i muscoli degli arti, lo sviluppo dei muscoli longitudinali del corpo, lo spostamento in un modo o nell'altro degli arti sotto il corpo - tutto ciò ha permesso di muoversi rapidamente con il corpo che si alza dal suolo (correndo). Il rafforzamento dei muscoli e il movimento in un ambiente diversificato è stato accompagnato da un aumento del carico sullo scheletro. Ciò ha richiesto il suo rafforzamento. Pertanto tessuto in fibra grossolana ossa tubolari trasformato in fibroso fine, le clavicole iniziarono a svolgere il ruolo di una specie di distanziatore tra le parti dorsale (bordo distale della scapola) e ventrale (sterno) della cintura della spalla. Il più comune era il tipo percentuale di vertebre. Tutte le vertebre del tronco, così come parte della cervicale e della caudale, portano costole mobili. Movimenti intensi di flessione - l'estensione della mano e del piede è diventata possibile a causa della comparsa di articolazioni intercarpali e intertarsali.
I rettili sono caratterizzati da una maggiore mobilità della testa. Ciò è garantito dall'articolazione del cranio estremamente mobile con la colonna vertebrale cervicale colonna vertebrale: in primo luogo, il cranio si collega alla prima vertebra cervicale con un condilo; in secondo luogo, le prime due vertebre cervicali (atlante ed epistrofia) vengono modificate in modo che siano possibili movimenti della testa attorno all'asse longitudinale del corpo. La mobilità della testa e del collo, combinata con la capacità di muoversi rapidamente attorno al substrato, offriva ai rettili ampie opportunità di trovare cibo. La forte compressione delle mascelle si è rivelata importante per catturare diverse prede. A questo proposito, si sono sviluppati potenti muscoli masticatori e sono stati ricostruiti il \u200b\u200btetto e i lati del cranio - sono comparse fosse temporali, che hanno aumentato significativamente la superficie per l'attacco dei muscoli della mascella. Deformazione del cranio sotto compressione muscoli da masticare impedito da ossa colonnari.
La connessione delle costole con lo sterno e la formazione del torace in questo modo è un'altra importante trasformazione dello scheletro dei rettili. Di conseguenza, il tasso di ventilazione dei polmoni è aumentato notevolmente e il volume di aria pompata è aumentato. Ciò ha contribuito al rifiuto della respirazione cutanea e all'aumento dell'intensità dello scambio di gas.
In generale, lo scheletro dei rettili porta un complesso di tratti che dimostrano un livello superiore di organizzazione di questi animali e adattamenti più profondi alla vita sulla terra.
Esplora lo scheletro dei rettili.
prendere in considerazione:
cranio
Cranio del cervello
occipitale: superiore; ossa occipitali principali e due laterali; forame occipitale; condilo occipitale.
Lati del cranio: malare; squamosa; palmati; una barra; square; ossa dell'orecchio.
Skull Roof: nasale; prefrontale; frontale; parietali; interparietal; strappo; fronte; le ossa postorbitali.
Fondo del cranio: osso sfenoide principale; apriscatole; rudimenti del palato osseo secondario.
Cranio viscerale
Mascella superiore: mascellare; ossa intermaxillari.
Mascella inferiore: dentale; articolare; angolazione; ossa arcuate;
Apparato sublinguale: osso ipoide con due paia di corna.
spina dorsale
La colonna vertebrale: collo; al seno; lombare; sacrale; coda; vertebra di tronco; Atlanta; Epistrophe. petto: sterno; costole.
Cintura degli arti
Cintura di spalla: sterno; torace; coracoid; scapole; clavicola.
Cintura pelvica: iliaca; sciatico; ossa pubiche; dell'acetabolo.
Arti accoppiati
forelimb: spalla; avambraccio (ulna e raggio); il polso; metacarpo; falangi a dita; articolazione intercarpale.
Arti posteriori: coscia; bacchetta (tibia e tibia); tarso; metatarso; falangi a dita; articolazione intertarsale.
Sketch:
1) cranio (vista dall'alto, vista laterale, fondo); 2) la cintura degli arti anteriori; 3) la cintura pelvica e la colonna vertebrale sacrale.
Struttura scheletro
Cranio del cervello
Nelle lucertole, il cranio è quasi completamente ossificato e consiste in un gran numero di ossa, formando un piuttosto massiccio scatola del cervello (Fig. 70, 71).
La regione occipitale del cranio consiste di quattro ossa occipitali, che limitano il forame occipitale. L'osso occipitale principale insieme a due ossa occipitali laterali forma il condilo occipitale, che entra nella fossa articolare dell'atlante. L'osso occipitale superiore a forma di arco copre il forame occipitale dall'alto ed entra dietro il tetto del cranio.
Lati del cranio nell'area della capsula uditiva sono formati da tre ossa dell'orecchio, che sono coinvolti nella formazione delle fosse temporali.
Fig. 70. La struttura del cranio di una lucertola:
la - vista dall'alto; B - vista dal basso; il - vista laterale; 1
- forame occipitale; 2
- osso occipitale laterale; 3
- osso occipitale superiore; 4
- il principale osso occipitale; 5
- condilo occipitale; 6
- l'osso sfenoide principale; 7
- apriscatole; 8
- cigni; 9
- parasfenoide; 10
- osso parietale; 11
- foro per l'organo parietale; 12
- osso frontale; 13
- osso nasale; 14
- osso intermaxillare; 15
- osso mascellare; 16
- l'osso prefrontale; 17
- osso lacrimale; 18
- ossa infraorbitali; 19
- osso postorbitale (posteriore); 20
- osso squamoso; 21
- osso temporale; 22
- osso zigomatico; 23
- narice; 24
- osso quadrato; 25
- osso palatino; 26
- osso pterigoideo; 27
- osso colonnare; 28
- osso trasversale; 29
- osso articolare; 30
- osso del dente; 31
- osso angolare; 32
- l'osso arcuato; 33
- osso coronarico

Fig. 71. La struttura del cranio di una lucertola monitor:
la - vista dall'alto B - vista dal basso 1
- osso intermaxillare, 2
- osso nasale 3
- l'osso prefrontale, 4
- osso frontale 5
- osso lacrimale 6
- osso infraorbitale 7
- osso parietale, 8
- osso occipitale superiore, 9
- il principale osso occipitale, 10
- osso occipitale laterale, 11
- osso squamoso 12
- osso quadrato 13
- osso postorbitale (posteriore), 14
- osso zigomatico, 75 - osso mascellare, 16
- apri 17
- osso palatino, 18
- osso trasversale 19
- osso pterigoideo 20
- principale sfenoide, 21
- forame occipitale, 22
- osso zigomatico quadrato, 23
- condilo occipitale
Le ossa anteriori sono ben sviluppate e isolate, le ossa uditive superiori si fondono con l'osso occipitale superiore e le ossa posteriori con le ossa occipitali laterali. Ai lati del cranio sono grandi orbite. Il setto interorbitale è formato da processi delle ossa frontali e parietali del tetto del cranio. Dietro l'orbita si trovano le ossa postorbitale, zigomatica e squamosa. Piccole ossa lacrimali si trovano nell'angolo anteriore dell'orbita e un certo numero di piccole ossa infraorbitali delimitano l'orbita dall'alto. La parte anteriore del cranio è formata da derivati \u200b\u200bdel cranio viscerale: ossa mascellari e intermaxillari. Nell'area olfattiva, la lucertola non ha ossificazione, la capsula olfattiva con le narici esterne è formata dalla cartilagine.
Tetto del cranio forma di ossa nasali, prefrontali, frontali e parietali accoppiate tra le ossa parietali si trova un osso inter-parietale non accoppiato con un'apertura per l'organo parietale.
Fondo del cranio formato dal principale sfenoidecon cui si fonde il rudimentale parasfenoide. Nella parte anteriore del parasfenoide sono le ossa palatine e gli apri, ai lati dei quali
ci sono hoans. Alcune ossa del cranio viscerale sono anche coinvolte nella formazione del fondo della scatola del cervello - queste sono palatine e ossa pterigoidi. Ossa pterigoidali curve con le estremità anteriori appoggiate contro le ossa palatine, e le estremità posteriori si articolano con lo sfenoide principale e con ossa quadrate.
Nella regione temporale delle lucertole si trova la fossa temporale, limitata da un singolo arco temporale superiore, che è formato dalle ossa postorbitale e squamosa. L'arco temporale inferiore nelle lucertole è ridotto. Nelle profondità della fossa temporale, le ossa colonnari (a forma di elitre) sono chiaramente visibili sotto forma di piccoli puntoni che poggiano sulle ossa pterigoidi e parietali.
Cranio viscerale
Arco mascellare nelle lucertole è fortemente modificato. Come risultato dell'ossificazione della cartilagine quadrata palatina, formarono ossa quadrate, pterigoidi, colonnari (alate elitre) e palatine. L'osso quadrato con la sua estremità superiore è articolato in modo mobile con un osso squamoso e quello inferiore con la mascella inferiore e funge da sospensione per esso. La comparsa di un'articolazione tra l'osso squamoso e l'estremità superiore dell'osso quadrato ci ha permesso di aumentare la forza di compressione delle mascelle e, apparentemente, ha causato il successo evolutivo delle lucertole. Mascella superiore formata dalle ossa mascellari e intermaxillari su cui si trovano i denti. Tra le ossa pterigoidi e mascellari si trovano le ossa trasversali, che fungono da distanziatori e rinforzano il cranio.
Base per lo sviluppo osseo mandibola serve cartilagine di meckel. È sostituito da un osso articolare articolato con un osso quadrato. Inoltre, la mascella inferiore comprende un numero di ossa tegumentarie: le più grandi ossa dentali che portano i denti e quelle più piccole - coronarie, angolari e arcuate.
Dipartimento superiore arco sublinguale - Iomandibolare - fa parte dell'orecchio medio sotto forma di un ossicolo uditivo - una staffa. Il resto dell'arco ipoide e i resti degli archi di branchia vengono convertiti nell'apparato ioidico, rappresentato dalla placca cartilaginea dell'ioide e tre coppie di corna.
spina dorsale
La colonna vertebrale dei rettili è divisa in quattro sezioni: cervicale, toracico, lombare, sacrale e caudale.
La maggior parte dei rettili ha vertebre protselnye, ad es. concavo anteriore. Il corpo vertebrale è allungato, è concavo di fronte,

Fig. 72. Le prime due vertebre cervicali di una lucertola monitor:
la - atlante; B - epistrophy; 1
- arco superiore; 2
- canale spinale; 3
4
- processo simile a un dente dell'epistrofia; 5
- corpo vertebrale
la duta superiore termina con il processo spinoso superiore. I processi articolari anteriori e posteriori si estendono dal corpo vertebrale. Alla base dell'arco superiore si trovano superfici articolate per il fissaggio di nervature.
Le prime due vertebre cervicali sono molto diverse dalle altre. La prima vertebra cervicale - atlante - ha la forma di un anello diviso da un fascio denso nelle metà superiore e inferiore (Fig. 72). Sulla superficie frontale dell'atlante è presente una fossa articolare per il condilo occipitale. Attraverso il foro superiore, il cervello si collega al midollo spinale e il processo simile a un dente della seconda vertebra cervicale entra nel foro inferiore nella parte posteriore - epistrofeya. Questa articolazione del cranio con il collo fornisce movimenti complessi della testa. Le costole dell'orso delle vertebre cervicali posteriori.
Alle vertebre toracica prendere parte costoletteche si articolano con lo sterno e formano la gabbia toracica (costole vere). Le vertebre lombari hanno anche costole che non si collegano allo sterno (false costole). Due vertebre sacrali hanno enormi processi trasversali che si articolano con la cintura pelvica (vedi Fig. 75). Il numero di vertebre caudali nelle lucertole varia, raggiungendo diverse decine. Le vertebre caudali anteriori portano processi trasversali e spinosi (Fig. 73) e, alla fine della coda, le vertebre vengono convertite in ossa a forma di bastoncino. I corpi di quasi tutte le vertebre caudali sono separati da un sottile strato cartilagineo, lungo il quale si verifica una rottura durante l'autotomia.

Fig. 73. La struttura della vertebra caudale anteriore della lucertola:
la - vista dall'alto; B - vista laterale; 1
- il corpo della vertebra; 2
- processo spinoso superiore; 3
- processo articolare anteriore; 4
- processo articolare posteriore; 5
- processo trasversale; 6
- processo spinoso inferiore; 7
- arco inferiore
Cintura degli arti
Cintura anteriore nelle lucertole (Fig. 74) è generalmente organizzato in modo simile alla cintura della spalla degli anfibi, tuttavia, rispetto ad esso, l'ossificazione è altamente sviluppata qui. Alla giunzione della coracoide con la scapola vi è una cavità articolare per l'articolazione omero. La cartilagine soprascapolare cresce fino al bordo dorsale della scapola e la cartilagine procoracoide cresce di fronte al coracoide. La coracoide e la procoracoide si fondono insieme, formando un singolo elemento scheletrico che si articola con lo sterno. In questa articolazione sono possibili movimenti scorrevoli su un piano orizzontale lungo il bordo anteriore dello sterno, ciò aumenta la lunghezza del passo. Dal basso allo sterno cresce un sovrasternale cruciforme di origine tegumentaria. Le clavicole collegano il bordo dorsale della scapola con il pettorale. Pertanto, a differenza degli anfibi, le clavicole dei rettili sono orientate su un piano verticale e rendono la cintura della spalla più resistente.
Cintura pelvica nelle lucertole è ben sviluppato e consiste di due ossa anonime, che sono state formate dalla fusione delle ossa pelviche: iliaca, sciatica e pubica (Fig. 75). Ci sono cavità articolari per la testa del femore (acetabolo). Le ossa pubiche e ischiatiche destra e sinistra sono collegate da una sinfisi cartilaginea. La cintura pelvica è saldamente collegata alla colonna vertebrale dalla crescita dell'ileo ai processi trasversali delle vertebre sacrali.
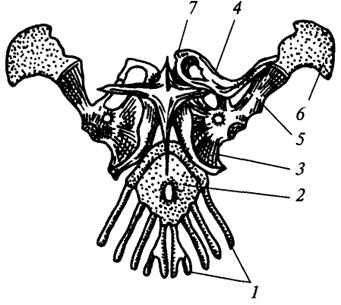
Fig. 74. La cintura di spalla di una lucertola:
1
- le prime quattro costole toraciche; 2
- sterno; 3
- coracoide; 4
- clavicola; 5
- scapola; 6
- cartilagine soprascapolare; 7
- cinghia pettorale

Fig. 75. Cintura pelvica di una lucertola: br 1
- cavità articolare (acetabolare) per la testa del femore; 2
- ilio; 3
- osso pubico; 4
- osso ischiatico; 5
- sinfisi
Arti accoppiati
Gli arti dei rettili corrispondono alla struttura generale dell'arto a cinque dita dei vertebrati terrestri. scheletro forelimb comprende omero, ossa ulnari e radiali, due file di piccole ossa del polso, una fila di ossa metacarpali e una fila di ossa delle falangi delle dita. scheletro arto posteriore include il femore, la tibia e la tibia, due file di piccole ossa tarsali, un certo numero di ossa metatarsali e una serie di falangi a dito. Le peculiarità della struttura delle estremità includono la formazione interkarpalnogo (nella parte anteriore) e tertarzalnogo invariante (nell'arto posteriore) delle articolazioni (Fig. 76).

Fig. 76. La struttura dell'articolazione intertarsale della lucertola (piede destro della lucertola veloce dall'alto):
1
- ossa tarsali prossimali fuse; 2
- ossa distali del tarso; 3
- tibia; 4
- piccola tibia; 5
- ossa del metatarso
L'articolazione intercarpale è formata tra due file di semi del polso, l'articolazione intertarsale è tra due file di ossa tarsali. Queste articolazioni offrono la possibilità di una forte flessione ed estensione della mano e del piede, il che è importante con un particolare tipo di locomozione.