Štruktúra lebky zvierat. Štruktúra mozgu zvierat a meningov. Mozgové tepny
SKELETON HEAD. SKULL
Vzhľad hlavy a jej kostry súvisel so komplikáciami celkovej organizácie predkov stavovcov, ktoré sa vyskytovali v boji o existenciu a vďaka postupnému vývoju tela, prispôsobujúcemu sa meniacim sa životným podmienkam. Na prednom konci bipolárneho tela zvieraťa sa vyvíjajú dve nezávislé časti kostry hlavy (obr. 32). Jedným z nich je ochranné zariadenie pre vyvíjajúci sa mozog - lebka - lebka, druhá (časť tváre) - v spojení so zlepšením zariadení pre lepšie a väčšie zachytávanie potravín a ich prvotné spracovanie. Na prednom konci tela dolných stavovcov sa objavili dve časti kostry hlavy: lebka, ktorá tvorí mozgovú lebečnú dutinu, a viscerálna časť, ktorá ďalej formuje tvár - mizne. Tieto dve oddelenia kombinovali všeobecnosť ich umiestnenia a skutočnosť, že mobilný čeľusťový prístroj časti tváre našiel podporu pre svoje svaly na lebke.
Mozog delfínov je druhým najsilnejším a najkomplexnejším mozgom u zvierat. Inteligenciu možno definovať ako mieru schopnosti mozgu spracovať informácie spôsobom, ktorý rieši problémy a zvyšuje prežitie. Meranie inteligencie môže byť úplne odlišné pre zvieratá, ktoré sa vyvinuli vo vode v porovnaní s tými, ktoré žijú na súši, pretože problémy potrebné na prežitie vo vode sa veľmi líšia od problémov vyžadovaných na súši. O „inteligencii delfínov“ sa veľa diskutuje.
Veľké mozgové zvieratá, ako sú ľudia, šimpanzy a delfíny, majú veľa spoločného, \u200b\u200bzvyčajne žijú dlhé životy, žijú v tekutých sociálnych skupinách av detstve vykazujú úplnú rodičovskú závislosť. Preto nie je možné úplne vyhodnotiť úroveň inteligencie tohto zvieraťa. Iným spôsobom, ktorý bol navrhnutý na meranie inteligencie, je stanovenie miery relatívnej veľkosti mozgu, ktorá je definovaná ako pomer medzi skutočnou hmotnosťou mozgu a predpokladanou hmotnosťou mozgu u zvieraťa danej veľkosti.
Obr. 32. Lebka žraloka
Každé oddelenie sa vyvinulo z rôznych prvkov nachádzajúcich sa v tejto oblasti: chrupavka vytvorená na stranách akordu a pred ním (lebka) a z prvých viscerálnych oblúkov (tvár). Kosti pochádzajúce z týchto miest podliehajú v ontogenéze trom stupňom osifikácie a sú sekundárne. Následne sa jednotlivé časti vonkajšej kostry, ktoré tvoria základné kosti lebky, podieľajú na tvorbe lebky. Krycie kosti lebky sú primárne a pri ontogenéze iba dva stupne osifikácie - membrána a kosť (obr. 33).
Toto meranie predpokladá, že čím vyššie číslo, tým vyššia inteligencia. Okrem toho sa stupeň koagulácie mozgovej kôry javí ako miera inteligencie. Čím väčšia je zložená kôra, tým viac priestoru v mozgu je možné umiestniť ďalšie neuróny, pomocou ktorých je možné vykonať spracovanie informácií. Túto teóriu podporuje nedávno publikovaná informácia o náraste skladania mozgovej kôry Alberta Einsteina v porovnaní s inými ľuďmi. Jediné zviera s komplexnejšou kôrou ako ľudia je delfín.
Vývoj stredného mozgu
Tento obrázok sa vzťahuje na ľudský mozog. hlavným dôvodom takého veľkého mozgu je spracovanie informácií z komplexnej echolokácie delfínov alebo biosonárnych systémov. Neexistuje však žiadny dôkaz na podporu tohto presvedčenia. Najbežnejšou teóriou je, že tento väčší mozog sa vyvinul na podporu komplexnejších kognitívnych schopností. Môžu si pamätať udalosti a študovať koncepty, ktoré menia svoje správanie v dôsledku predchádzajúcich skúseností. Môžu komunikovať medzi sebou počas kooperatívneho správania, riadiť vzťahy vo svojich tobolkách a vychovávať svoje deti.
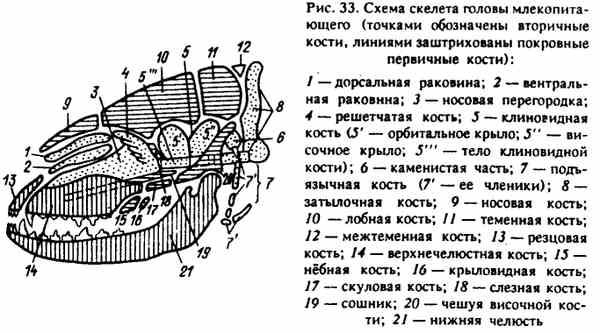
Obr. 33. Schéma kostry hlavy cicavca (bodky označujú sekundárne kosti, línie pokrývajú základné primárne kosti)
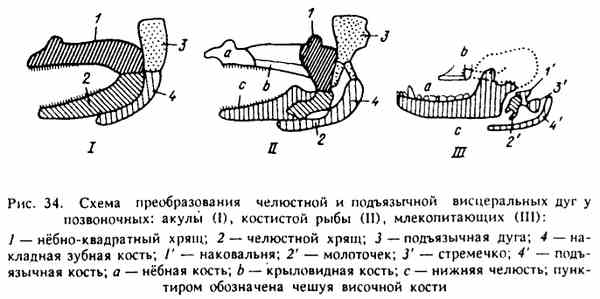
Obr. 34. Schéma transformácie čeľustných a sublingválnych viscerálnych oblúkov na stavovcoch
Dokážu porozumieť nielen slovám symbolického jazyka, ale tiež interpretujú syntaktické poradie jazyka. Toto porozumenie syntaxe veľmi svedčí o inteligencii. Pohyblivé píšťalky vytvorené delfínmi sú dôkazom toho, že delfíny majú sebavedomie alebo schopnosť mať koncept seba a vedieť, že existuje ako individuálna bytosť. Sebapovedomenie existuje v prednej čelnej kôre. Okrem delfínov existuje očividne sebapoznanie iba u veľkých mozgových primátov a ľudí.
V dôsledku štúdia Alzheimerovej choroby u ľudí sa zistilo, že neuróny ľudského vretienka sú spojené so schopnosťou rozpoznávať, zapamätať si, rozumieť, komunikovať, vnímať, prispôsobovať sa zmenám, riešiť problémy a porozumieť. Ohniská pacientov s Alzheimerovou chorobou vykazujú vysoký výskyt opotrebovaných vretenových neurónov. Nedávno boli neuróny vretena izolované v mozgu skutočných veľrýb. Tento záver ukazuje, že skutočné veľryby majú tieto pokročilé schopnosti, ktoré sa v minulosti spájali iba s ľuďmi a primátmi.
Lebka bola vytvorená z mnohých kostí, ktoré tvorili samostatné časti lebky a tváre. Medzi kosťami sú švy, ktoré sú vyplnené hustou spojivové tkanivo, Medzi niektorými kosťami v ontogenéze sa tvoria významné spojivové tkanivové priestory nazývané fontanely; do vysokého veku sú osifikované. U cicavcov sa teda takéto fontanely nachádzajú medzi týlnymi a parietálnymi kosťami lebky alebo medzi týlnymi, parietálnymi a temporálnymi.
Všeobecné charakteristiky vývoja mozgu
Už dlho existuje podozrenie, že delfíny majú schopnosť rozoznať ľudí a predmety, zapamätať si úlohy, riešiť problémy, prispôsobiť sa zmenám a študovať zložité úlohy. Inštitút delfínov v Honolulu na Havaji skúma správanie morských cicavcov za posledných 30 rokov. Prostredníctvom svojej práce značne rozšírili naše znalosti správania delfínov a inteligencie. Nasleduje stručný prehľad výsledkov niekoľkých výskumných projektov.
Všeobecné vlastnosti štruktúry mozgu
Delfíny, rovnako ako ľudia, sú schopné prejavov správania v tvári. Inými slovami, môžu napodobňovať správanie demonštrované ich ľudskými trénermi. Ak človek zdvihne nohu, delfín dokáže rozoznať vzťah, ktorý má časť ľudského tela pre svoju vlastnú anatómiu, a zdvihne chvost. To naznačuje, že zviera môže asociovať časť svojej anatómie s ľudskou formou.
V priebehu fylogenézy niektoré kosti, ktoré tvoria kostru hlavy dolných stavovcov, následne menia svoje funkcie. Primitívne čeľuste rýb sú teda nahradené klasickými kosťami, ale čeľusťové kosti vôbec nezmiznú, menia svoju funkciu, objavujú sa na horných stavovcoch zvukovými kosťami stredného ucha (obr. 34).
U suchozemských zvierat sa v súvislosti so zmenou dýchania žiabrov na pľúcne dýchanie vyvíja nosová dutina na báze čuchových fosílií na tvári, ktorá v prednej časti komunikuje s úzkym otvorom v ústach medzi rezákom, maxilárnymi kosťami a otváračom. Tieto otvory sa nazývali primárne hoany. U zvierat s primárnymi choanmi nemôže potrava prejsť cez ústnu dutinu, keď vzduch prechádza nosovou dutinou. V tomto ohľade sa štruktúra nosovej dutiny zlepšuje a komplikuje. Poloha chób sa postupne mení a už u plazov sa otvára za nosnú dutinu do hltanu a nazýva sa sekundárnymi chónami.
Môže napodobniť aj iného delfína. Dôkazom toho je výkon synchrónneho správania. Ak sa jeden delfín chystá pokloniť, iné zviera môže toto správanie skopírovať a súčasne s ním skákať. Hovorí sa, že jeden delfín pôsobí ako „demonštrant“, zatiaľ čo iné zviera je „mimikom“ tohto správania.
Delfíny dokážu interpretovať aj televízne správanie a reagovať na gestá, ktoré sa zobrazujú na obrazovke po prvom vystavení televízoru. Toto je prvá demonštrácia akejkoľvek reakcie zvierat na správanie pri televíznych gestách. Psy, mačky a šimpanzy takéto reakcie nevykazovali v podobných výskumných protokoloch. Táto štúdia zmenila, ako sme v minulosti klasifikovali delfíny predovšetkým ako akustických špecialistov. Teraz chápeme, že sú tiež vizuálnymi špecialistami a využívajú vizuálny aj zvukový výkon, aby vynikali vo svojom vodnom prostredí.
Lebka je spojená s prvým krčným stavcom (atlasom) a tvorí lebečnú dutinu, v ktorej sa nachádza mozog. Táto lebečná dutina - cavum cranii má strechu, zadné, predné, bočné steny a spodok lebečnej dutiny.
SKULL STRECH - kalvária.
U ošípaných, koní a psov je tvorený párovou prednou kosťou - front frontale (pred), párovou parietálnou kosťou - os parietale a nepárovou inter-dark dark - os interparietale- (zozadu). Parietálna kosť kaudálne hraničí s týlnou kosťou. U mladých zvierat sa v mieste stehov medzi parietálnymi a týlnymi kosťami vytvára týlny fonanel, v ktorom sa položí párová osifikácia osifikácie, z ktorej sa vytvorí nepárová medzipastická kosť. Nachádza sa v strede línie v rohu medzi dvoma parietálnymi a týlnymi kosťami. Z vnútornej strany lebečnej dutiny je možné vidieť výrazné zhrubnutie kosti, z ktorého vystupujú hrebene doprava a doľava. Toto je test na kosti - tentorium osseum, ku ktorému je pripojený membránový test dura mater.
Ukázalo sa, že delfíny sa v zrkadle rozpoznávajú pomocou tzv. „Kontingenčného testovania“ alebo uskutočňovaním pohybov a skúmajú tento pohyb. Tento objav je neočakávaný, pretože delfíny primárne prežívajú svet prostredníctvom zvuku a ich echolokačný systém nebude fungovať v dvojrozmernej reflexii.
Koniec mozgu - telencefalon
Sú si vedomí svojho nedávneho správania a môžu opakovať správanie alebo, ak o to tréner požiada, vykonať správanie, ktoré sa nedávno nevykonalo. Družstvá predstavujúce „opakovanie“ alebo tímy predstavujúce „akékoľvek“ majú za následok opakovanie nedávneho správania alebo výber iného nedávneho správania. Ukazuje to schopnosť delfínov udržiavať mentálne správanie, ktoré vykonal naposledy, a aktualizovať tento obraz pri vykonávaní každého nového správania, pričom na požiadanie opakuje posledné správanie v tejto sekvencii.
U koní, ošípaných a psov je predná časť strechy lebečnej dutiny tvorená frontálnym čelným sklom. Je to parný kúpeľ, reprezentovaný tromi doskami:
1) horný predný, ku ktorému je nosová kosť pripevnená vpredu;
2) orbitálne, zostupne bočné a zúčastňujúce sa spolu s krídlami sfenoidnej kosti a šupín časná kosť vo formácii mimo lebky vnútornej steny obežnej dráhy. (Na križovatke týchto dvoch doštičiek odchádza zygomatický proces - procesový zygomatik, ktorý u koní a prežúvavcov dosahuje zygomatický proces časnej kosti alebo dočasný proces zygomatickej kosti a tvorí úplný kruh obežnej dráhy u ošípaných a psov, odchádza mimo prednej kosti na styčnej ploche týchto dvoch doštičiek.) nedosahuje a tvorí neúplný prsteň. Na základe tohto procesu nájdete infraorbitálne foramen - foramen supraorbitály.
Delfíny reagujú na trénera, ktorý ukazuje na objekt. Delfíny nielen rozumejú a primerane reagujú na osobu, ktorá smeruje priamo na objekt, ale zodpovedajú aj na priesečník. Príkladom vhodnej reakcie na indikáciu by bolo získanie objektu, na ktorý školiteľ odkazuje, alebo presunutie objektu z bodu A do bodu B.
Podobná štúdia schopnosti delfínov zostať zameraná na úlohy vyžadujúce dlhodobú koncentráciu priniesla zaujímavé výsledky. To dokazuje schopnosť delfínov zostať na pozore po dlhú dobu a rýchlo rozlišovať medzi kritickými a nekritickými obrázkami a zvukmi s vysokou mierou presnosti.
U prežúvavcov sa supraorbitálny foramen nachádza na začiatku zygomatického procesu, z ktorého sa priekopy pohybujú tam a späť a prechádzajú na obežnú dráhu do kanála.
U ošípaných prechádza diera do žlabu a dosahuje nosné kosti.
Psi nemajú infraorbitálne foramen.);
3) vnútorná vertikálna (nie je zvonka viditeľná), ktorá tvorí prednú časť strechy lebečnej dutiny.
Okrem sebauvedomenia delfíny prežívajú základné emócie, podieľajú sa na určitom stupni abstraktného, \u200b\u200bkoncepčného myslenia, vyberajú si svoje činy, učia sa pozorovaním, rozumejú štruktúre svojho prostredia, učia sa, čo funguje a čo nie, riešením problémov. vytvárať nové riešenia problémov, s ktorými sa stretávajú. Zdá sa, že pri interakcii s osobou sú si vedomí rozdielu medzi deťmi a dospelými a majú tendenciu byť jemnejší a trpezlivejší s deťmi.
Neuroveda naznačuje, že ľudský mozog je štandard, podľa ktorého by sa mali porovnávať mozgy všetkých zvierat. Mnoho zvierat však vykazuje jedinečný talent a vysoko pokročilé mentálne schopnosti a spoločenské správanie. Ich inteligencia je veľmi odlišná od človeka a nejakým spôsobom ho prekonáva. Mnohé výskumné oblasti ukazujú jedinečné živočíšne mozgy s jedinečnými schopnosťami obnovy zvierat.
Medzi týmito tromi doskami (prednými, okružnými a vnútornými) sa v pravej a ľavej prednej kosti tvoria medzery - dutiny vyplnené vzduchom a pokryté sliznicami. Nazývajú sa dutiny alebo dutiny. V prednej kosti je to spárovaný čelný sínus - sínusový frontalis. Symetrické dutiny sú oddelené sagitálnym septom a nie sú komunikované. Vo vnútri dutín sú stále nespojité oddiely, ktoré rozdeľujú každý sínus na bunky, ktoré spolu komunikujú. Čelné dutiny prechádzajú do parietálneho a týlneho (u ošípaných a hovädzieho dobytka).
Slávny filozofický článok sa pýtal, či dokážeme pochopiť, aké to je byť netopierom. V skutočnosti je nemožné, aby sa ľudia dostali do skúsenosti bytostí s úplne odlišnými zmyslovými a zmyslovými vlastnosťami pohonné systémy, Štruktúry v mozgu zvierat sú úplne odlišné od ľudí a hodnotenie ich funkcie v porovnaní s ľudským mozgom nevysvetľuje veľmi pokročilé správanie.
Väčšina zvierat má oveľa viac inteligencie, ako by sa dalo očakávať jednoduchým anatomickým porovnaním s ľudským mozgom. V súčasnosti je dobre známa inteligencia slonov, delfínov, veľryb, chobotníc a primátov. Nedávno sa zistilo, že delfíny si spomínajú na spev svojich tankistov spred dvadsiatich rokov - neuveriteľná pamäť. Pokročilá inteligencia terapeutických psov bola objavom toho, že mnoho zvierat má kultúru.
U prežúvavcov je výraznou črtou lebky to, že iba čelná kosť tvorí strechu lebečnej dutiny.
U hovädzieho dobytka má významné rozmery, vo vnútri dosť silných dutín, prechádzajúcich do parietálnych a nosných dutín a stále dutých nadržaných procesov - processus coretimees, ktorých dutina komunikuje s čelnými dutinami, odchýliť sa od vonkajšej plochy. Preto si musíme pamätať na to, že pri rezaní rohu sa vstup do predného sínusu otvára otvorom vytvoreným v procese dutého rohu prednej kosti. Zadná časť lebečnej klenby tvorí zosilnenú hranu (vyvýšenie medzi rohmi), ku ktorej sú pod ňou pripevnené kosti parietálnej a medziparietálnej. Je to podobné týlu na iných zvieratách. Čelná kosť, ktorá tvorí celú strechu lebečnej dutiny u hovädzieho dobytka, akoby tlačila parietálne a intertmavé kosti, ktoré sa u týchto zvierat nachádzajú za, mierne nad týlnou kosťou, pod čelným hrebeňom, k zadnej a bočnej stene dutiny. Parietálne kosti vstupujú do bočnej steny medzi predné (nad) a dočasné (pod) kosti, získavajú tvar písmena P, na ktorého hornej časti čelná kosť, od spodnej časti (za) týlnej kosti, a po stranách (pod) dočasné kosti.
Žilové dutiny mozgu
Tento príspevok bude venovaný niekoľkým mozgom mozgu s odlišnou štruktúrou mozgu ako ľuďom, ale výnimočným skautom, plazom a včelám. Existuje mnoho ďalších možných príkladov veľmi malých zvierat s inteligenciou, ako sú mravce a termiti, ale medzi hmyzom boli najviac študované včely. Správa uzatvára rady o tom, ako môžu tieto malé mozgy mať pokročilé funkcie s mozgovými štruktúrami, ktoré sa líšia od ľudí.
Vtáky potrebujú pomerne veľký mozog, aby ich veľkosť mohla letieť. Ale v porovnaní s ľuďmi a inými veľkými zvieratami sú ich mozgy veľmi malé. Nedávno sa zistilo, že kakadu majú mimoriadnu schopnosť používať viacstupňové metódy na odomknutie veľmi zložitej bunky. Na odstránenie matice vták najskôr odstránil kolík, potom odstránil skrutku, potom odstránil skrutku, potom otočil koleso o 90 stupňov a potom zatlačil západku na stranu. Tieto vtáky neboli pripravené a dokázali prísť na to, ako to urobiť za menej ako dve hodiny.
U oviec sa na rozdiel od hovädzieho dobytka časť parietálnych kostí podieľa na tvorbe zadnej časti strechy lebečnej dutiny a smerom k týlnej oblasti tvorí hrebeň parietálnej kosti.
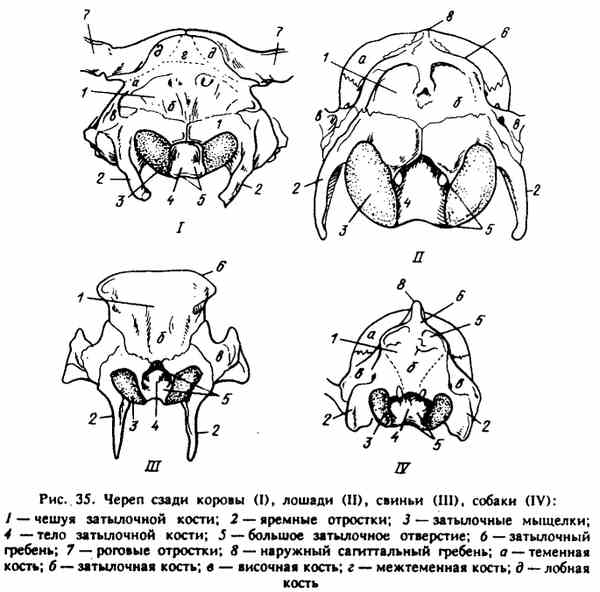
Obr. 35. Lebka za kravou (I), koňom (II), prasa (III), psom (IV)
Mozgové tepny
Ostatní sa mohli učiť sledovaním. Akonáhle sa im podarilo otvoriť jeden zo zámkov, nikdy nezabudli, ako by to mohli urobiť okamžite. Okrem toho bolo nedávno kakadu úplne vyvrátené myšlienkou, že zvieratá nemôžu tancovať, a okrem toho si všímajú stálosť objektu na úrovni štvorročnej osoby alebo opice.
Jace, spolu s papagájmi, spevmi vtákov a havranmi, patria medzi najchytrejších vtákov. Nedávno sa zaznamenalo, že počas smútku zosnulého kamaráta počas 48 hodín bolo bez kŕmenia pozorované celé kŕdeľ sôp. Kaledónsky havran mal vynikajúcu pamäť, vrátane spomienok na ľudí a autá, ktoré ich o mnoho rokov neskôr obťažovali. Preukazujú metacognition a počítanie, a tiež robiť a používať pokročilé nástroje lepšie ako mnoho primátov. Inovovali nástroje, vytvarovali kúsok drôtu do háčika, aby získali potešenie z ťažko dosiahnuteľného miesta.
Pre kosti lebky je charakteristické, že na vnútornej strane kraniálnej dutiny majú kosti hladký povrch, na ktorom sú viditeľné malé priehlbiny - zahĺbenia, tzv. Odtlačky prstov - digitatae. Toto sú mozgové výtlačky.
ZADNÁ STENA KRÁLNEJ ÚROVNE (Obr. 35). Tvorí týlnu kosť - týl. Má tri ohniská enchondrálnej osifikácie, preto sa u mladých zvierat delí na tri časti: šupiny - squamaoccipitalis, condylar (laterálne) časti - partes laterales a telo týlnej kosti (hlavná časť) - pars basilaris. Bočné (condylar) časti obklopujú veľký otvor - foramen magnum na výstup z miechy do miechového kanála. Pri silnom naklonení hlavy nadol medzi týlnu kosť a atlas sa vytvára priestor - prístup k mieche, ktorá sa používa pri zabíjaní zvierat. Na stranách veľkého otvoru sú zaoblené condyle týlnej kosti, condyli occipitales, ktoré sú pokryté kĺbovou chrupavkou, ktoré spolu s kĺbovými otvormi atlantátu tvoria atlantokokový kĺb.
Bočné kondyly sú krčné procesy - procesné jugulary. Sú veľmi silné, rovné u ošípaných, o niečo menšie u koní a prežúvavcov a malé, mierne zaoblené u psov.
Na základe jugulárnych procesov leží mediálne okrúhle hyoidné otvorenie - foramen hypoglossi, ktoré prechádza do hyoidného kanála, cez ktorý posledný - dvanásty pár kraniálnych nervov - hyoidný glosár - vychádza z lebečnej škatule.
U dospelého zvieraťa sa fúzujú všetky tri časti týlnej kosti. Váhy týlnej kosti u koní, ošípaných a psov sú umiestnené nad kondylom a jej horná časť tvorí silný týlny (outdent) hrebeň - crista occipitalis nuche). Má trojuholníkový tvar a tvorí ošípaný obdĺžnik.
SÚŤAŽ KRÁĽOVSKEJ KRIVKY (Obr. 36). Tvorí ju hlavná časť (telo) týlnej kosti a sfénoidnej kosti. Pozdĺž okraja tela lícnej kosti - foramen lacerum sa vytvára členitá diera (niekedy vo forme medzery u ošípaných a koní), cez ktorú vystupujú hlavové nervy (vetvy V, IX, X a XI).
Sphenoidná kosť - os sphenoidale umiestnená pred spodnou časťou lebky. Ak ho izolujete od lebky (v mladom veku, až kým sa kosti neuzavrú vo švíkoch), vyzerá to ako motýľ. Jeho stredná / nepárová časť - telo - corpus sphenoidale sa nachádza pred telom týlnej kosti. Pozdĺž bokov tela vedú dva páry tenkých kostných dosiek. Predné sú o niečo väčšie ako zadné - krídla sfenoidnej kosti - alae sphenoidaly, spolu s telomgóniou tvoria predné a stredné lebečné kosti, v ktorých leží stredný a terminálny mozog.
Venujte pozornosť vnútornému povrchu tela sfénoidnej kosti, má dva kroky: horná predná časť je presfenoidná, zadná a spodná sú základným fenotypom, ktoré sú od seba oddelené úzkou priečnou medzerou, ktorá sa nazýva črevo vizuálneho prieniku - sulcus chiasmatis. U mladých zvierat je medzi nimi synchoda ruží. V dolnom kroku - fundamentenoid, je vidieť, že fosíliu prechádza do eminencie zozadu (na hranici tela tela). Tieto útvary pripomínajú tvar ázijského sedla, v súvislosti s ktorým dostali názov „turecké sedlo“ - sella turcica. Vo svojej fosílii leží hypofýza (endokrinná žľaza), pred ktorou je vo žľabe vizuálneho prieniku priesečník optických nervov, ktoré prenášajú impulz z sietnice oka do mozgu.
Na prednom okraji vonkajšieho povrchu krídel sféenoidnej kosti sú umiestnené otvory zhora nadol, cez ktoré kraniálne nervy spájajú mozog s orgánmi hlavy umiestnenými na prednej časti steny obežnej dráhy. Najvyšší je etmoidný otvor - foramen ethmoidale (tvorí ho aj predná kosť), pod ním je optický canalis canalis opticum, pod ktorým kôň a pes majú orbitálnu fisúru - fissura orbitalis a dokonca nižšie kruhový otvor - foramen rotundum (u ošípaných a prežúvavcov) kombinované do okrúhleho otvoru).
Na vonkajšej strane sfenoidnej kosti sú umiestnené pterygoidné procesyrámovanie obruby. Sú veľké a chromé u hovädzieho dobytka a ošípaných. U koní a psov prechádza pterygoidným kanálom canalis pterygoideus, do ktorého prechádza maxilárna tepna a nerv, základom týchto procesov.
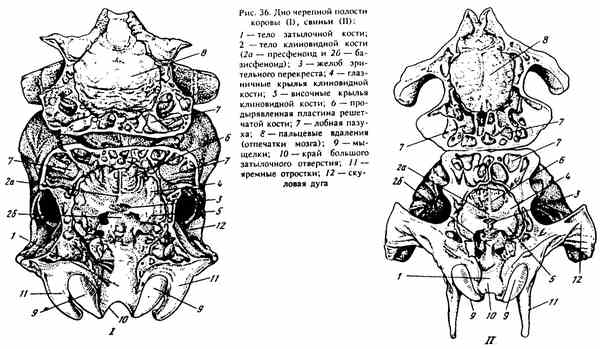
Obr. 36. Spodok lebečnej dutiny kravy (I), ošípané (II)
Bočné steny stenovej jaskyne. Zhora sú pravé a ľavé párové krídla sfenoidnej kosti spojené švami so šupinami spánkových a čelných kostí a tvoria bočné steny lebečnej dutiny. V mladom veku je tu neosifikovaná oblasť - klinovitý fontanel - fonticulus sphenoidal.
Dočasná kostná os je zapojená do tvorby bočnej steny kraniálnej dutiny. Má štyri časti: šupinatá časť - pars squamosa - tanier, ktorý tvorí bočnú stenu; kamenná časť - pars petrosa - v kaudálnej časti lebečnej dutiny; časť bubna - pars tympanica - na vonkajšej strane lebky, vo vnútri ktorej vyniká vnútorná časť bubna - pars endotympanica.
Zygomatický proces temporálnej kosti, processus zygomatics os temporale, sa bočne vetví od vonkajšieho povrchu spánkových šupín, ktorý sa ohýba vpred a spája sa s časovým procesom zygomatickej kosti, pričom s ňou tvorí zygomatický oblúk - arkus zygomaticus. Na ventrálnom povrchu lamelárneho zygomatického procesu je v segmentovej rovine kĺbový povrch na spojenie s dolnou čeľusťou.
Vo svojej prednej časti sú časové stupnice a krídla sfenoidné kosti tvoria bočné steny na vnútornej strane lebečnej dutiny a na vonkajšej strane stien obežnej dráhy, na povrchu ktorých sa otvára etmoid, optický kanál, orbitálna puklina a kruhový otvor, ktorým prechádza päť lebečných nervov (páry II, III, IV, VI) a vetvy V páru), v dôsledku čoho je táto časť spodnej časti obežnej dráhy jednou z najsytenších kraniálnych nervov.
Kaudálna časť časných kostných šupín pokrýva kamennú hruď - pars petrosa (Obr. 37), ktorá sa tiež podieľa na tvorbe bočnej steny kraniálnej dutiny a leží pred a nad krčkovými procesmi. Táto časť je najviac „nezávislá“ v prípade (je relatívne ľahké ju oddeliť), u ostatných zvierat je veľmi pevne spojená s váhami. Skalnatá časť je určená predovšetkým na umiestnenie sluchových a rovnovážnych orgánov. Je viditeľný zo strany lebečnej dutiny, rovný, hladký, nesúci odtlačky stočených mozgov. Vo vnútri je uložený kostný labyrint - kostné dutiny rôznych tvarov, v ktorých sú umiestnené orgány sluchu a rovnováhy. Z tohto labyrintu vychádzajú na povrch kamennej časti dve malé trhliny - vonkajšie otvory kochleárneho kanála a predsieňový apertura externa vestibuli a apertura externa aqueductus cochlea.
Toto treba pamätať na to, že práve cez ne dutina kostného labyrintu vnútorného ucha komunikuje s intershell priestormi lebečnej dutiny; ochorenie načúvacích orgánov môže v tomto ohľade viesť k ochoreniu meninges - meningitídy.
Na boku lebečnej dutiny v strednej časti povrchu kamennej časti je pomerne veľká priehlbina, z ktorej vedú dve kosti: vnútorný zvukový mušus - muškátový vývar, ktorým je nervový systém (VIII) spojený s drieňovou oblongatou nervom (VIII); otvor pre tvárový nerv (VII), ktorý spája mozog s tvárovými svalmi.
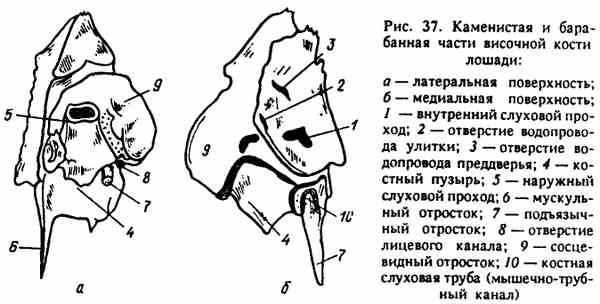
Obr. 37. Kamenné a tympanické časti časnej kosti koňa
Na vonkajšej strane kamennej časti lebky sa nachádza tympanická časť, pars tympanica, a nad ňou sa nachádza mastoidný proces, processus mastoideus. Na strane bubna je zreteľne viditeľná významná (najmä u prežúvavcov a ošípaných) bubnová bublina - bulla tympanica. Vo vnútri je dutina - cavum tympani (poloha stredného ucha), do ktorej vedie špičková kostná trubica veľmi významného priemeru. Táto trubica je vonkajším zvukovým mäsom - meatus acusticus externus, vodivým zvukom, ktorý putuje z vonkajšieho ucha cez stred k vnútornému. Na spodnej časti vonkajšieho zvukovodu je zreteľne viditeľný proces hystid (styloid) - procesus hyoideus (stylordeus) (najmä u koňa), ktorý sa vyznačuje výskytom naostrenej valcovej kostičky. Hyoidná kosť - hyoideum je pripojená k hyoidnému procesu, na ktorom je zavesený jazyk, hltan a hrtan. Tu môžete vidieť ostrý svalový proces, stiahnutý, ku ktorému sú pripojené svaly palatínovej opony. Na jeho spodnej časti je viditeľná veľká medzera, ktorá vedie tiež k dutine bubienkovej bubliny, kde je umiestnené stredné ucho. Táto medzera je svalová trubica (zvuková trubica) - canalis muscu-lotubarius (tuba audiva), cez ktorú stredné ucho komunikuje s hltanovou dutinou. Toto je cesta infekcie z hltanu do stredného ucha.
Mastoidný proces - pocessus mastoideus je pomerne rozsiahly, vyrobený z špongiová látkapokryté tenkou vrstvou výliskov. Podľa neho osoba používa načúvací prístroj v prípade, že je vylúčená možnosť viesť zvukovú vlnu cez stredné ucho.
Pod mastoidným procesom sa otvára štrk - mastoid - foramen styJomastoideum, ktorým miechový nerv (VII) opúšťa kamennú časť, ktorá inervuje tvárové svaly.
Dno kamenistej časti na hranici tela telesa kosti sa môže podieľať na tvorbe členitej diery.
PREDNÁ STRANA KRÁSNEJ CAVITY. Pred sphenoidnou kosťou, ktorá tvorí prednú stenu lebečnej dutiny, sa nachádza etmoidná kosť - os ethmoidale. Táto kosť nie je viditeľná na povrchu lebky. Leží na hranici medzi lebečnými a nosnými dutinami. Z vnútornej strany prednej steny lebečnej dutiny je možné vidieť dve zaoblené vybrania, na spodnej časti ktorých sú viditeľné dosky s hromadou dier, jedná sa o perforovanú dosku - lamina cribrosa. V dierach tejto doštičky prechádzajú nervy do cibuliek čuchového mozgu, ktoré spájajú mozog s čuchovým systémom nosovej dutiny.
Zo strany nosovej dutiny je k perforovanej doske pripevnená hmota malých, rôznych veľkostí najtenších kostných dosiek tvoriacich kučery. Tieto vonkajšie a vnútorné turbíny sú endoturbinálne a ektoturbinálne. Celkovo tvoria tieto kučery labyrint - labyrint ethmoidalis. Pravá a ľavá časť labyrintu sú oddelené pomerne tenkou doskou umiestnenou pozdĺž stredovej sagitálnej línie, ktorá sa nazýva kolmá doska. Z boku lebečnej dutiny je jej začiatok viditeľný vo forme zvisle nasadenej skalpely umiestnenej medzi dvoma vybraniami perforovanej platne.
Z boku nosovej dutiny pokračuje kolmá doska kostí etmoidnej kosti vpred vo forme už chrupavkovitej, nikdy neosifikujúcej nazálnej septa. Z kolmej doštičky zo strany nosovej dutiny napravo a naľavo sú po stranách tenké kosti zvlnené doštičky obklopujúce labyrint. Ukazuje sa, že hlavná časť, etmoidná kosť, labyrint, v ktorom sa nachádza čuchový orgán, leží medzi tromi doštičkami, ktoré tvoria druh „škatule“ s dvoma oddeleniami (táto škatuľa je rozdelená do dvoch symetrických oddelení kolmou doskou). Zadná stena škatule (dierovaná doska) je viditeľná z boku lebečnej dutiny. Z boku nosovej dutiny je upevnený labyrint. Tento box je otvorený iba zo strany nosovej dutiny.
Aby ste dobre porozumeli umiestneniu a názvu všetkých dier, ktorými mozog ležiaci v lebečnej dutine komunikuje s orgánmi, do ktorých jeho nervy idú, skúste sa znova pozrieť na tieto dierky na lebke. Na prednej stene cez otvory perforovanej platne etmoidnej kosti komunikuje kraniálna dutina s labyrintom etmoidnej kosti, t.j. s nosnou dutinou. Na zadnej stene, cez veľký (týlny) otvor, mozog prechádza do miechy a kraniálna dutina komunikuje s miechovým kanálom. Na stranách spodnej časti lebky tvorenej telami týlnych a sfenoidných kostí sa nachádza séria dier, cez ktoré kraniálne nervy spájajú mozog s rôznymi orgánmi hlavy a tela. Na vonkajšom povrchu prednej časti steny obežnej dráhy prechádza zhora nadol reťaz vertikálne usporiadaných otvorov: etmoidný, optický kanál, orbitálna puklina a okrúhla, trochu roztrhaná diera, viac kaudálna pozdĺž tela týlnej kosti, za ktorým je okrúhly sublingválny otvor na strane mediálneho povrchu jugulárnej procesnej základne ( sublingválny nervový kanál).
Mozgové škrupiny
Žilové dutiny mozgu
Mozgové tepny
Všeobecné vlastnosti štruktúry mozgu
Všeobecné charakteristiky vývoja mozgu
Koniec mozgu - telencefalon
Vývoj mozgu
Stredná mozog-Diencephalon
vývoj medzimozgu
Stredná mozog-Mesencephalon
Vývoj stredného mozgu
Zadný mozog-Rhombencephalon
Vývoj kosoštvorcov
Zoznam použitej literatúry:
Mozgové škrupiny
Dura mater - dura mater encephali - najviac vonkajšie. Na rozdiel od tvrdého obalu miechy rastie spolu s periosteom kraniálnych kostí, takže tu nie je epidurálny priestor. Medzi periosteom a dura materom prechádzajú iba žily, ktoré tvoria dva systémy žilových dutín - dorzálnu a ventrálnu.
Do spodného priestoru od dura mater sa rozprestierajú dva záhyby: kosákovitý a membránový obrys mozgu. Kosiakovité kosáky v tvare kosiaka sagitálne prechádzajú od hrebeňa kohútika etmoidnej kosti k mozočku. Leží v pozdĺžnej medzere medzi mozgovými hemisférami. Najviac kosáčikovitý záhyb sa prejavuje u psov a koní - dosahuje takmer corpus callosum; je menej u ošípaných a kráv, najmä u malých prežúvavcov. Membránový cerebelárny stan - tentorium cerebella membranaceum - sa odchyľuje od kostnej značky alebo parietálnej kosti, ako aj od hrebeňov. kamenné kosti a nachádza sa v priečnej medzere medzi veľkým mozgom a mozočkom.
Arachnoidálna membrána mozgu arachnoidea encephali - na zvraty mozgu pevne spojené s mäkkou škrupinou. Subarachnoidálny priestor je zachovaný iba v štrbinách a ryhách medzi zvlneniami a na bazálnom povrchu mozgu, kde vytvára nadstavby - ventrálne cisterny drene oblongata, cisternový most, červ (a iné). Pod arachnoidálnou membránou pozdĺž kosáčikového záhybu idú villi do subdurálneho priestoru, niekedy vo forme uzlov - pachyónových granúl - granulácií arachnoidálov. Pachyónové granule sa zavádzajú do sagitálneho sínusu a zvyšujú odtok mozgomiechového moku do žíl.
Mäkká alebo choroidná pia materská encefalia - veľmi pevne spojený s mozgom, prechádza do všetkých trhlín a výklenkov a spolu s cievami preniká do mozgu. Keď sa otočí späť do dutiny mozgu, podieľa sa na tvorbe cievnych pneumatík - tela chorioidea -, v ktorých sú umiestnené cievne plexusy-plexus chorioideus. Takéto útvary sa nachádzajú v komorách mozgu: bočné, tretie a štvrté.
Cerebrospinálny likér s tekutým likérom - sú vyplnené subdurálne a subarachnoidálne priestory mozgu a miechy a cez párové otvory - apertura lateralis ventricularis kvartakudal z laterálnych nôh mozočku - a nie spárovaný otvor - apertura mediana ventriculi kvarti komunikuje s dutinami komôr a mozgových kanálov. Cerebrospinálna tekutina je sekretovaná bunkami ependymy a vaskulárneho plexu. Tečie v subarachnoidálnom priestore smerom k mozgu, tj kraniálne a kaudálne v centrálnom kanáli miechy. Výtok mozgomiechového moku sa vyskytuje z subdurálneho priestoru do žilového systému a zo subarachnoidálneho priestoru do lymfatických orgánov.
Žilové dutiny mozgu
Dorsálny sínusový systém pozostáva z nepárového sagitálu a priameho sínusu a spárovaného korenia, týlneho a dorzálneho kameňa; do nich prúdia žily mozgu.
Sagitálna sínusová dutina sagitálna je umiestnená v kosáčikovom záhybe; nazálne to začína z membránových žíl, u koňa tiež z etmoidných a mozgových žíl. Do toho spadajú:
1) dorzálne žily mozgu-vv.cerebri dorsales;
2) membránové žily z dura mater;
3) kostné žily-vv. diploicae;
4) krátky priamy sínusový sínusový konečník.
Ten je tvorený kaudálne z corpus callosum fúziou žíl corpus callosum s veľkou mozgovou žilou.
Corpus callosum vein-v. corporis callosi - nesie krv z prednej polovice mozgu.
Veľká žila mozgu-v. cerebri magna-berie do hlbokých žíl mozgu-vv. cerebri, ktorý odoberá krv z mozgu a jej vaskulárnych plexov.
Kaudálne z sútoku priameho sínusu je sagitálny sínus rozdelený na pravý a ľavý priečny sínus, -sinus transversus - každý z nich je zameraný na dočasný kanál a prechádza do dorzálnej mozgovej žily-v. cerebri dorsalis. Ten prúdi do časovej povrchovej žily. Kamenný sínusový sínusový petrosus, ktorý nesie krv zo základných častí mozgu, tečie do priečneho sínusu predtým, ako sa ponorí do dočasného kanála.
Medzi priečnymi dutinami sú spojovací sínusovo-sínusoví komunikanti - leží na spodku mozočkového stanu. Do nej spadnú týlne dutiny, ktoré sa nachádzajú v brázdach na obidvoch stranách mozočka.
K odtoku krvi z dorzálneho systému dutín dochádza okrem mozgových dorzálnych žíl tiež prostredníctvom emissáriového emissária do dočasných hlbokých žíl. Emiséri sú nekonzistentné vypúšťacie kanály, ktoré perforujú stenu lebečnej dutiny.
Ventrálny alebo bazilárny sínusový systém zahŕňa sínusový kruhový a bazilárny.
Kruhový sínusový sínusový kruh obklopuje hypofýzu. Tvoria ho pravé a ľavé kavernózne dutiny, ktoré spájajú medzistavcové dutiny - sinusové kavernózne a interkaverózové.
Každý kavernózny sínus nazálne prechádza do orbitálnej žily mozgu kaudálne do bazálneho sínusu. Okružná žila mozgu-v. cerebralis orbitalis - tečie orbitálnou puklinou do hlbokej žílnej tváre.
Bazilárna sinus sinus basilaris - infúzia komorových stavcov. V oblasti členitej diery dáva ventrálnu a mozgovú žilu-v. cerebralis ventralis - ktorý tečie do týlnej žily. V blízkosti týlneho kĺbu tvoria bazilárne dutiny, medzi ktorými sú anastomózy, ventrálny týlní dutín sínusový occipitalis ventralis. Ventrálne žily mozgu sa otvárajú do bazálneho sínusového systému.
Mozgové tepny
1. Vnútorná krčná tepna-a. Carotis interna - preniká do lebečnej dutiny cez karotický alebo roztrhnutý otvor a je rozdelený na nazálne a kaudálne spojovacie vetvy. S vetvami toho istého mena na druhej strane tvoria okolo mozgového arteriálneho krúžku-cirkula arteriózy. Nosná tepna-a vychádza z prstenca nazálne. cerebri nasalis - ide do corpus callosum a vyživuje predné hemisféry.
4 nosné vetvy z nosnej spojivovej vetvy:
a) nazálna artéria meninges-a. meningea nasalis;
b) stredná mozgová artéria-a. cerebri media prechádza na bočný povrch mozgu;
c) nazálna artéria vaskulárneho plexu-a. chorioidea nasalis - pozdĺž optického traktu sleduje vaskulárny plexus laterálnej komory;
d) vnútorná orbitálna tepna-a. oftalmika interna- tuje cez optický nerv na obežnú dráhu.
Od kaudálnej spojovacej vetvy odídte:
a) kaudálna cerebrálna artéria-a. cerebri caudalis - pre zadné časti pologule a štvornásobok
b) kaudálna artéria vaskulárneho plexu-a. chorioidea caudalis-v choroidálnom plexe laterálnych komôr.
2. Z týlnej tepny miechová tepna-a odchádza. cerebrospinalis - cez medzistavcové otvorenie atlasu preniká do stavcovského kanála a je rozdelený na kraniálne a kaudálne vetvy, pričom na druhej strane spája rovnaké vetvy. Hlavná tepna mozgu je oddelená od lebečných vetiev v mozgu. basilaris cerebri- tečie do arteriálneho krúžku a na svojej ceste dáva: za mostom je kaudálna tepna mozgu, pred mostom je lebečná tepna mozočka. cerebella caudalis et cranialis - a do sluchového nervu - vnútorná sluchová tepna - a. auditiva interna. Kaudálna vetva anastomózy miechy s miechovou ventrálnou tepnou-a. spinalis ventralis. Mozgové cievy sú inervované sympatickým nervovým systémom.
Všeobecné vlastnosti štruktúry mozgu
Mozog - encefalon - z dorzálneho povrchu je rozdelený priečnou fisúrou - fissura transversa cerebri - na veľký a kosoštvorcový mozog.
Mozog sa skladá z dvoch hemisfér, diencefalonu a stredného mozgu. Pravá a ľavá hemisféra mozgu - hemisphaerum cerebri dextrum et sinistrum - sú dorzálne oddelené hlbokou pozdĺžnou puklinou, fissura longiscere cerebri. Medziprodukt a stredná mozgu sú dorzálne zakryté hemisférami.
Rhomboidphalon-rhomboidný mozog sa skladá z mozgu, mozgového mosta, kombinovaného do zadného mozgu, a medully oblongata.
Cerebellum cerebellum je umiestnené dorzálne od drene oblongata a zadne po mozgovej hemisfére.
Medulla oblongata-medulla je priamym pokračovaním miechy. Na prednom konci je most cerebri-pons cerebri.
Pred mostom sú nohy veľkého mosta. V prednej časti sa nohy rozprestierajú k optickým traktom - tractus opticus - a optickým chiasm-chiasma opticum. Medzi nohami veľkého mozgu, za vizuálnym prierezom, ležia: mastoidné telo-korpus mamillare- a sivý tubercle s lievikom a hypofýzou. Pred optickými ústrojenstvami a po stranách nôh mozgu vyčnieva čuchová mozgová rinencefalon, ústne končí dvoma čuchovými žiarovkami, bulbi olfactorii. Čuchový mozog predstavuje ventrálnu časť mozgových hemisfér a maskovanú paládium-dorso-laterálnu časť pokrytú brázdami a stočeniami.
Hemisféry patria do terminálneho mozgového telencefalonu - a do optických traktov, sivého tuberkulu, lievika, hypofýzy a mastoidov - do ventrálnych častí diencephalonového diencefalonu.
Nohy veľkého mozgu sú ventrálnou časťou mezenecephalonového stredného mozgu.
Mozoček a most sú časťou metamorfónu hindubra. Medziprodukt, meduľa a stredná mozog sa kombinujú do kmeňa mozgovo-črevného mozgu.
Všeobecné charakteristiky vývoja mozgu
Mozog sa vyvíja v súvislosti so vznikom a ďalším vývojom rôznych vyšších nervové centrá: receptor, motorický a asociatívny, ktoré zabezpečujú koordinovanú funkciu všetkých orgánov - somatických, viscerálnych, cievnych.
V skorých štádiách ontogenézy a fylogenézy je mozog zväčšený koniec mozgovej trubice, ktorá leží pred akordom, a preto sa nazýva predchordálny alebo primárny mozgový mechúr. Jeho vývoj je spojený s čuchovým orgánom a jeho čuchová funkcia je zachovaná u všetkých zvierat vrátane cicavcov.
O niečo neskôr je na prednom konci epichordálneho mozgu izolovaný predchordálny alebo sekundárny mozgový mechúr. Jeho vývoj je spôsobený: vznikom primárneho asociatívneho a komisárskeho centra; prítomnosť žiabrových prístrojov a orgánov laterálnej línie vodných živočíchov; diferenciácia vnútorných orgánov.
Z prednej časti epichordálneho mozgového mechúra sa oddeľuje stredný mozgový mechúr, ktorý s výnimkou cicavcov zohráva vedúcu úlohu, pretože impulzy prichádzajú rozhodujúcim spôsobom zo všetkých receptorových aparátov, najmä vizuálnych.
Z chrbtového povrchu sú všetky tri časti mozgu celkom ostro oddelené od seba priečnymi komisiami nervových vlákien, pred a za strednou mozgovou vrstvou.
Následne sa predchordálny mozog rozdelí na terminálny mozog vo forme párového mechúra a diencefalonu.
Terminálny mozog dosahuje najvyššie štádium vývoja u cicavcov s prítomnosťou novej mozgovej kôry, ktorá sa stáva vyšším nervovým centrom.
V diencephalone u nižších zvierat je v súvislosti s vedúcou úlohou analyzátora čuchovcov na prvom mieste hypotalamus a epithalamus. Sú to stredné čuchové centrá, ktoré vysielajú impulzy do stredného mozgu a hypotalamus navyše vysiela impulzy do hypofýzy a do autonómnych stredov stredných a stredných oblongat. Z tohto dôvodu sa hypotalamus stáva najvyšším subkortikálnym vegetatívnym centrom. Chráni tiež túto funkciu u cicavcov.
Iba s tvorbou mozgovej kôry u cicavcov sa intenzívne vyvíja stredná časť diencefalonu, talamus, prostredníctvom ktorého kôra rozhodne prijíma všetky receptorové impulzy z periférie.
Preto je hypotalamus historicky viac asociovaný s epichordálnym mozgom a thalamus s predchordalom.
Takmer súčasne s rozdelením predchordálneho mozgu sa epichordálny mozog diferencuje za strednou mozgu na zadnú mozgu a medulla oblongata. Zadný mozog pre cicavce predstavuje iba mozoček, ktorý je subkortikálnym centrom korelácie svalových pohybov na udržanie rovnováhy. Iba u cicavcov je mozgový mostík pridaný do mozgu v dôsledku vývoja nového mozgového kôru a samotné mozog je posilnené dvoma mozgovými hemisférami.
Dutiny primárnych mozgových mechúrov sa stávajú mozgovými komorami v rozvinutom mozgu. Z dutiny predchordálneho mozgu v terminálnom mozgu vznikajú párové bočné komory a v diencephalone tretia mozgová komora. Všetky tri komory sú spojené interventrikulárnym otvorom. Komora stredného mozgového mechúra v dôsledku rastu stien poslednej transformácie na mozgový akvadukt a kosoštvorcová dutina sa stáva štvrtou mozgovou komorou, ktorá komunikuje s centrálnym miechovým kanálom.
Koniec mozgu - telencefalon
Skladá sa z dvoch mozgových hemisfér - hemisphaerum dextrum et sinistrum - oddelených od dorzálneho povrchu hlbokou pozdĺžnou puklinou - fissura podélná cerebri.
Na každej pologuli sa skúma plášť, čuchový mozog, striatum a laterálne komory mozgu. Plášť sa nachádza v hemisfére dorzálne od srdcovej komory, čuchový mozog je komorový. Hranica medzi plášťom a čuchovým mozgom na ventrálnom povrchu mozgu je základná hraničná drážka. Striatum leží vo ventrálnej stene hemisféry dorzálne z častí čuchového mozgu.
A) Plášťové paládium
Pozostáva zo sivej a bielej mozgovej hmoty. Šedá meduľa-substantia grisea- tvorí na svojom povrchu mozgovú kôru - mozgovú kôru. Na ňom rozlíšiť zvraty mozgového vírusu - oddelené ryhami a trhlinami. Na bočnom povrchu plášťa sú viditeľné tri klenuté gyusy. Obklopujú laterálny sylvian sulcus-sulcus lateralis. U psov sú zreteľne výrazné oblúky a u ostatných zvierat je každý oblúk rozdelený na časti zárezmi. Na strednom povrchu plášťa sú viditeľné hlavne dva oblúkovité gyrusy okolo tela corpus callosum, pričom najbližší cingulate gyrus girus cinguli- funkčne spojený s čuchovým mozgom.
Pod kôrou plášťa plášťa sa nachádza biela drieň substantia alba-mask. Pozostáva z ciest - asociatívne, komisariálne a projekčné.
a) asociatívne vlákna spájajú samostatné časti kôry v každej hemisfére. Rozdeľujú sa na krátke a dlhé vlákna.
b) komisurálne vlákna spájajú oblasti patriace do rôznych hemisfér. Tvoria corpus callosum corpus corpus, najväčší komisár mozgu. Je umiestnený medzi hemisférami v hĺbke pozdĺžnej medzery. Odlíšte kmeň tela corpus callosi truncus corporis corporis a dva konce - predné a zadné. Predný koniec sa nazýva koleno corpus callosum-true corporis callosi - ohýba sa ventrálne. Zadný koniec alebo vankúšik corpus callosisplenium corporis corpus je spojený s oblúkom.
Commissurálne vlákna, ktoré vychádzajú z kmeňa corpus callosum, vytvárajú žiarenie komissus-radiatio corporis callosi-, tvoria dorzálnu stenu laterálnej srdcovej komory mozgu a rozbiehajú sa do prednej, bočnej a zadnej časti plášťovej kôry.
c) projekčné vlákna - spájajú kôru plášťa s jednotlivými časťami mozgového kmeňa a s miecha, Tvoria vnútornú kapsulu v striate. Funkčne premietacie dráhy sú rozdelené na efferentné aferentné.
Eferentné dráhy prenášajú impulzy z kortexu mozgových hemisfér do rôznych častí mozgového kmeňa: veľké, kosoštvorcové a miechy.
Aferentné dráhy prinášajú impulzy do mozgovej kôry z miechy a kosoštvorca. Tvoria zadnú časť vnútornej kapsuly.
Na rozdiel od mozgovej kôry, všetky sivé látky zostávajúcich častí celého stredu nervový systém zjednotený pod pojmom „subkortex“. Impulzy zo všetkých častí tela idú najskôr do rôznych častí subkortexu, vrátane vizuálnej tuberkulózy a z druhej vstúpia do kortexu hemisféry. Kôra neustále rozpadá, rozpadá excitácie vznikajúce v receptorových zariadeniach a súčasne kombinuje, syntetizuje účinok excitácií izolovaných analýzou do rôznych komplexov. Toto je prejav analytickej a syntetickej aktivity mozgovej kôry. Najvyššia časť nervového systému - mozgová kôra (u ľudí) je navyše materiálnym substrátom vyššej nervovej aktivity: myslenie, vedomie, pamäť, rozum, vôľa, to znamená všetka duševná aktivita, ktorá je výsledkom fyziologickej aktivity mozgovej kôry.
B) čuchový mozgovo-rinencefalon
Nachádza sa v ventrikulárnej časti každej hemisféry veľkého mozgu. Niektoré jeho časti sú viditeľné na bazálnych a stredných povrchoch hemisfér, ako aj na spodnej časti laterálnych komôr mozgu. Čuchové žiarovky, čuchové ústrojenstvo a gyrus, čuchové trojuholníky a hruškovité laloky sa nachádzajú na spodnom povrchu hemisfér. Na stredných povrchoch hemisfér sú viditeľné čuchové pole, hippocampus gyrus, cingulate gyrus a plocha na narezanie nosovej adhézie a na spodku laterálnych komôr mozgu sú viditeľné jadrá kaudátu, hippocampus a oblúk. Bedrový gyrus, hippocampus, oblúk a amygdala sú limbický systém-pars limbica rhinencephalon.
1. Cibuľová cibuľka-cibuľka olfactorius - párová formácia vo forme pomerne plochého, pretiahnutého a zakriveného dorzálneho mozgového procesu, ktorý vyčnieva za prednú hranu hemisféry mozgu do čuchovej dutiny etmoidnej kosti. Dorsomediálna časť žiarovky je postavená z šedej drene a laterálne - ventrálne - z bielej drene. V žiarovke sa nachádza komora čuchovej banky - ventriculus bulbi olfactorii - je to pokračovanie laterálnej komory mozgu. Čuchová cibuľka obsahuje čuchový nerv-n. olfactorius. Obsahuje množstvo zväzkov nervových vlákien - fila olfactoria - nasmerovaných z čuchových buniek nosovej sliznice do nervových buniek cibule. Čuchové cibuľky sú teda primárnymi čuchovými centrami.
2. Od nervové bunky čuchové žiarovky Začnú čuchové cesty , Tvoria bielu medullu samotnej cibule a čuchových traktov - spoločné, stredné a bočné traktus olfactorius communis, medialis et lateralis. Bočný čuchový trakt prechádza do hruškovitého laloku, ktorý pokrýva bočný čuchový gyrus girus olfactorius lateralis po celej svojej ceste. Stredný čuchový trakt dosahuje stredný povrch plášťa a vytvára parolfaktórnu plochu poľa. Čuchové úseky obmedzujú čuchový trojuholník - trigonum olfactorium - zo sivej hmoty. Vedú impulzy z čuchovej cibule do buniek sekundárnych čuchových centier v čuchovom gyruse, čuchových trojuholníkoch, čuchových poliach, v hruškovitých lalokoch, ako aj v hypotalame a v strednom mozgu.
3. Lobus periformis v tvare hrušky - umiestnené stredne od bočného čuchového traktu a kaudálne od čuchového trojuholníka; stredne ohraničené nohami veľkého mozgu. Kaudomediálna hranica hruškovitého laloku je strednou medzerou alebo fissura hippocampi. Dutina je uzavretá v hruškovitom laloku, ktorý je zadnou časťou laterálnej srdcovej komory mozgu.
Hruška v tvare hrušky kaudálne prechádza bez jasnej hranice do gyrusu hippocampu, ktorý sa nachádza na strednom povrchu hemisféry, zozadu a laterálne od medzery hippocampu.
Hippocampus gyrus dorzálne pokračuje do cingulate gyrus girus cinguli. Ten prechádza priamo dorzálne z tela corpus callosum a ohýba sa okolo neho vpredu, pripája sa na čuchové pole a je súčasťou kôry plášťa. Hruška v tvare hrušky je sekundárnym čuchovým centrom.
4.Hippocampus alebo ammonský roh-hippocampus - svojou dorzálnou časťou tvorí spodnú časť laterálnej komory mozgu, za jadrom kaudátu, od ktorej je oddelený vaskulárny plexus laterálnej komory. Hippocampus je záhyb mozgovej kôry v oblasti hippocampálnej pukliny a laloku. Polmesiac sa ohýba laterálne 0-kaudálne a ventrálne a stráca sa v stene v tvare hrušky. Ammonské rohy ležia chrbtovo na vizuálnych tuberkulózach a sú od nich oddelené vaskulárnym plexom tretej mozgovej komory. Ako najvyššie asociatívne subkortikálne čuchové a chuťové centrá sú amoniakové rohy spojené s rôznymi časťami mozgovej kôry a subkortikálnych jadier. Ich vodivé cesty tvoria trezor a jeho deriváty.
5. Sejf - fornix - obsahuje cesty spájajúce hippocampus s mastoidným telom diencephalonu. Oddelené časti tohto zväzku vlákien tvoria drážkovanú vrstvu, okraj amónnych rohov, nohy, stĺpy, telo a prednú komisiu mozgu.
Drážkový listový alveus hippocampi pokrýva hippocampus tak, aby jeho povrch smeroval k laterálnej mozgovej komore. Tvoria ho nervové vlákna pochádzajúce zo šedej drene hruškovitého laloku a hippocampu. Pozdĺž dorsolaterálneho okraja tieto vlákna vytvárajú hranicu hippocampus-fimbria hippocampi-, ktorá sa nazálne rozprestiera do končatiny arch-crus fornicis. Ten, ktorý sa na druhej strane spája s nohou, sa stáva krátkym telesom arch-corpus fornicis. Telo oblúka slúži ako chrbtová stena tretej mozgovej komory; nazálne sa delí na dva stĺpce arch-columnae fornicis. Posledne menované stredne od jadier kaudátu sú smerované k mastoidnému telu a šedému knollu hypotalamu. Commissure archu - comissura fornicis - je tvorená priečnymi vláknami medzi nohami oblúka; spája chrbtové konce amónnych rohov.
6. Medzi predným koncom tela corpus callosum a stĺpmi oblúka je predná komisia mozgu-comissura nasalis. Spája čuchový mozog oboch hemisfér medzi sebou.
C) Striatum corpus striatum
Nachádza sa pred vizuálnymi tuberkami, dorzálne od čuchových trojuholníkov, pozostáva zo štyroch jadier: kaudátu, lentikúl, mandlí a plotu. Medzi jadrámi sú vnútorné a vonkajšie adhézie kapsuly tvorené projekčnými cestami.
Na jadre kaudátu sa rozlišuje jadro kaudatus, hlava a chvost. Hlava caudátového jadra s dorzálnym povrchom tvorí spodok laterálnej komory, naso laterálne od hippocampu.
Lentikulárne jadro, nucleus lentiformis, je umiestnené laterálne od hlavy caudátového jadra a optického tuberkulózy a je oddelené internou internou kapsulou-kapsulami. Bočná časť lentikulárneho jadra sa nazýva škrupina putamen. Stredná časť lentikulárneho jadra staršieho pôvodu sa nazýva bledé jadro-globus pallidus. Je spojený s čuchovým mozgom, leží laterálne od optického tuberkulu a pred jadrom bočnej kľukatej gangliovej genikulatum. Vonkajšia vonkajšia kapsula-kapsula je umiestnená laterálne od lentikulárneho jadra a druhé jadro, laterálne od kapsuly, je druhým jadrom vo forme úzkeho pruhu mesta claustrum. Na hranici medzi plotom, škrupinou a amónnym rohom sa nachádza jadro amygdaly, jadro amigdale, ktoré je čuchovým mozgom.
Pruhované telieska sú spojené cestami: 1) s mozgovou kôrou; 2) s vizuálnymi tuberkulózami a hypotalamom; 3) s jadrom retikulárnej formácie stredného mozgu, s jadrom mostíka a drôtenou oblongata; 4) s jadrom hlavových nervov.
Prostredníctvom pruhovaných telies sú rôzne reflexné reťaze uzavreté alebo:
A) prístroje na periférne receptory - vizuálne hľuzy - striatum - prístroje na somatické a viscerálne efektory.
B) kortex - striatum - somatické a viscerálne efektorové prístroje.
U cicavcov sú jadrá striata najdôležitejšími subkortikálnymi motorickými centrami:
1) koordinované nedobrovoľné hnutia;
2) Regulácia svalového tonusu v stave pokoja a pohybu;
3) nepodmienené reflexy;
4) Vyššie subkortikálne vegetatívne centrá
Pruhované telá fungujú ako celok, ale ich jednotlivé časti pôsobia opačne.
Vývoj mozgu
Zo starších predných a ventrálnych stien terminálneho mozgu pochádzajú čuchové laloky a striatum az jeho neskoršej dorzálnej steny - plášť.
1. Vývoj čuchových lalokov mozgu je determinovaný prítomnosťou analyzátora čuchov, ktorý je u vodných živočíchov mimoriadne dôležitý pri orientácii vo vonkajšom prostredí. Procesy citlivých čuchových buniek končia čuchovými žiarovkami, ktoré tvoria časť čuchových lalokov terminálneho mozgu, ktorých proximálne úseky tvoria najstaršiu čuchovú kôru, paleokortex, zo šedej medully. U cicavcov tvorí čuchový gyrus a čuchové trojuholníky - trigonum olfactorium - čo sú sekundárne čuchové centrá.
2. Vo ventrálnej stene terminálneho mozgu, dorzálne od čuchovej kôry, sa veľmi skoro objaví bazálna ganglio-gangliová bazalka veľkých buniek. Toto je najvyššie motorové centrum. Bazálny ganglion sa u cicavcov zachováva ako bledé jadro. Neskôr u suchozemských zvierat rastú ďalšie malé bunkové jadrá, ktoré tvoria celé jadro, ktoré pozostáva z lastúry, a u cicavcov jadro kaudátu. Okrem toho sa izolujú nové jadrá - plot a amygdala. S príchodom sekundárneho plášťa je komplexný bazálny ganglion prepichnutý vnútornými a vonkajšími kapsulami z ciest, ktoré prechádzajú do kôry plášťa, a naopak. Na základe toho sa objavil nový pojem - pruhované telo.
3. Pri vývoji plášťa sa pozorujú dve formácie rôznych funkcií a štruktúr, ktoré nepočítajú membránový primitívny plášť-pallium membranaceum - charakteristický pre vodné živočíchy a pozostávajúci z ependymy. U suchozemských zvierat sa najskôr objaví kôra jednoduchšej trojvrstvovej bunkovej štruktúry. Z toho sú tvorené vyššie čuchové centrá: hruškovité laloky, gyrus hippocampu a ich deriváty. Všetky tieto časti sú zjednotené konceptom primárneho plášťového arhipaléria.
Primárny plášť je charakterizovaný umiestnením sivej drene pod bielou driekom. Archipálium, ako aj paleopálium, sa najlepšie prejavuje v makrozmatických zvieratách - makrosmatesoch - s akútnym zápachom, na rozdiel od mikrosmatmatikov, v ktorých je čuch menej rozvinutý.
U cicavcov sa na hranici medzi čuchovým lalokom a primárnym plášťom objaví sekundárny neopáliový plášť. Sekundárny plášť sa vyznačuje povrchovým usporiadaním sivej drene vo forme kôry nad bielym drieňom. Základy sa objavujú u plazov. Ako rastie, nový plášť je tlačený z Arichipalluimu na strednú stranu a ventrálne. Súčasne sa v oblasti hippocampálnej pukliny vytvára záhyb kôry vo forme hippocampu a samotné neopálium zaberá celú dorsolaterálnu časť hemisféry a pri šírení kaudálne pokrýva strednú a dokonca strednú mozgovú vrstvu z ich dorzálnych a laterálnych povrchov.
Rast sekundárneho plášťa je spôsobený zavedením nových projekčných nervových vlákien z diencephalonu, ktoré sú vodičmi rôznych analyzátorov - kožných, zrakových, zvukových, svalov a vývoja nových asociatívnych neurónov a centier vrátane retikulárnej formácie.
Kôra sekundárneho plášťa v sérii zvierat je svojou štruktúrou mimoriadne zložitá, jej funkcie a štruktúra sú výrazne rozlíšené a jeho veľkosť sa zvyšuje. U veľkých cicavcov má plášť obvykle gyrus a ryhy. Takéto zvieratá sa nazývajú gyrencefali. Meandre v sérii zvierat nie sú rovnaké. V niektorých prípadoch ide hlavne o oblúky okolo priečnej sylvianskej drážky; u primátov tvoria gyrusy dva systémy - frontálny a parietálny. Oba systémy sú oddelené Sylvianskou brázdou. V tretej skupine zvierat sylvická bráza chýba a brázdy prebiehajú pred mozgom pozdĺžne a priečne vzadu. Homologizácia konvolúcií medzi zvieratami rôznych rádov je preto mimoriadne náročná av niektorých prípadoch to môže byť nemožné.
U malých zvierat nie je gyrus zvyčajne prítomný. Najväčší počet súžení sa vyskytuje u slonov a veľrýb. Pri ontogenéze sa gyrus tiež neobjavuje okamžite, ale v určitom poradí.
Stredná mozog-Diencephalon
Nachádza sa za striatom terminálneho mozgu a pred midbrainom, dorzálne zakrytým vaskulárnou výstelkou tretej mozgovej komory a hippocampu. Diencephalon sa skladá z troch oddelení rôzneho pôvodu, štruktúry a funkcie: epithalamus, talamus a hypotalamus. Epitalamus je tvorený vaskulárnou pneumatikou tretej mozgovej komory, epifýzou a párovým frenumálnym uzlom. Talamus sa skladá z vizuálnych tuberkulóz, medzi ktorými je tretia srdcová komora v tvare prstenca. Hypotalamus pozostáva z vizuálneho výčnelku s koncovou doskou, sivého tuberkulínu s lievikom a hypofýzou a mastoidného tela. Všetky časti hypotalamu sú viditeľné na bazálnom povrchu mozgu, medzi nohami veľkého mozgu, za priesečníkom optických nervov.
Talamu. Optické tuberkulózy - talamie - najmasívnejšia časť diencephalonu. Naso-laterálne sa spájajú s kaudátovými jadrami striata; vizuálne hľuzy budú od nich oddelené hraničným pruhom - stria terminis -, od kvadrupólu priečnou drážkou a od seba fossou vizuálnych hľúz, zakryté vaskulárnou výstelkou tretej komory. Rany tvoria početné zhluky jadier šedej drene. Najväčší z nich:
1.nasal nucleus-nucleus nasalis thalami - leží v hrúbke nosového tuberkulóza-tuberkulum nasalis thalami- v nasoediálnej časti optického tuberkulózy. Je to najstaršia a je centrom prechodu aferentných trás na reflexné cesty.
2. Caudal nucleus-nucleus caudalis thalami - uzavreté v hrúbke kaudo-laterálnej časti laterálneho tuberkulózneho tuberkula lateralis thalami- a pozostávajú zo stredných vizuálnych a zvukových stredísk. Z priesečníka optických nervov a sluchových centier. Od priesečníka optických nervov, chiasma opticum, na bazálnom povrchu mozgu začínajú optické trakty tractus opticum. Každý trakt sa ohýba bočne okolo talamu a prechádza do laterálneho ohnutého tela, corpus geniculatum lateralis, ktoré sa stráca v kaudálnom jadre vizuálnych ciest, ktoré prechádzajú do mozgovej kôry. Medzi bočným zalomeným telom a štvornásobkom vyčnieva mediálny zalomený korpus geniculatum medialis. Spája kaudálne kopce štvorkolky s kaudálnym jadrom optického tuberkulu a je stredným zvukovým centrom na ceste do kôry. Obidve kľukové telá sa kombinujú do 7.
3. thalami laterálneho jadra-jadra lateralis - je centrom na prepínanie vodivých dráh analyzátora kože a analyzátora pohybového aparátu na dráhy vedúce k mozgovej kôre a striatu.
4. stredné jadro-jadro medialis thalami - je stredným motorickým strediskom pre cesty od kôry k jadru extrapyramidálny systém: do červeného jadra, do jadier kraniálnych nervov a do miechy.
5. čistá tvorba-formatio reticularis - umiestnené medzi jadrámi a spojené s nimi.
Tretia komora mozgu --ventriculus terius - sa nachádza medzi optickými tubermilami, má kruhový tvar, pretože do neho rastie stredná hmota optických tuberkúl. V stenách komory je stredná sivá meduľa-substantia grisea centralis - obsahuje subkortikálne autonómne centrá. Tretia komora komunikuje s mozgovým akvaduktom stredného mozgu a za nosovou komisiou mozgu - comissura nasalis - s laterálnymi komorami mozgu prostredníctvom interventrikulárnych foramen interventriculare.
epithalamus , Na okrajoch fossy vizuálnych tuberkulóz sú viditeľné mozgové pásy vizuálnych tuberkulóz - stria medullaris - a na nich je párový uzol gangliovej habenuly. Uzol lievika prechádza do lýtka alebo vodítka-habenula, čím sa posilňuje tvar epifýzy - epifýzy - hrušky. Šišinka je endokrinná žľaza, ležiaca vo fossa medzi vizuálnymi tuberkulózami a štvornásobkami. Ganglion frenula slúži ako stredné centrum pre reflexné dráhy medzi mozgom, jadrom piateho páru a intersticiálnym jadrom.
Vaskulárna výstelka tretieho mozgu srdcovej komory chorioidea ventriculi terii - tvorená záhybom epitelovej platne mäkkej membrány mozgu a vaskulárneho plexu. Epitelové platne pneumatiky sú pripevnené pozdĺž okrajov fossa vizuálnych tuberkúl a oblúka. Vaskulárna pneumatika oddeľuje optické tuberkulózy od amoniakových rohov a od oblúka, preniká interventrikulárnym otvorom do laterálnych komôr mozgu vo forme vaskulárnych plexov laterálnych komôr mozgu-plexus chorioideus ventriculi lateralis. Vaskulárny obal sa tvorí pred epifýzou a bezprostredne za corpus callosum cusp-recus suprapinealis.
hypotalamus - subtalamický, starodávnejší čiastočný hypotalamus - tvorí stenu tretej srdcovej komory ventrálne od vizuálnych hľúz, je najvyšším subkortikálnym vegetatívnym centrom, spája sa s vizuálnymi tuberkulózami, s čuchovým mozgom as midbrainom. Hypotalamus obsahuje centrá regulácie: dýchanie, krvný obeh, teplota, metabolizmus, sexuálne funkcie. Prednú časť predstavuje šedý tubercle, lievik a hypofýza, zadnú časť telo mastoidu a steny tretej komory.
Hypotalamické hľuzy cinereum - leží priamo za priesečníkom optických nervov medzi nohami veľkého mozgu. V strede šedého kopca sa nachádza zátoka infundibuli z nálevky-výtoku. Samotný lievik - infundibulum je tenkostenné, k nemu je pripojená hypofýza.
Hypofýza - je plocho-kruhové teleso zložitej štruktúry s malou strednou dutinou. Hypofýza sa skladá z troch častí: mozgovej, strednej a žľazy. Je to najdôležitejšia žľaza s vnútornou sekréciou, ktorá vylučuje rôzne hormóny a reguluje všetky ostatné žľazy s vnútornou sekréciou, sympatický a parasympatický nervový systém.
Mastoidný corpus mamillare - leží priamo za šedým tuberkom a slúži ako stredné reflexné čuchové centrum, ktoré je prostredníctvom komplexu oblúkových útvarov spojené s čuchovým mozgom. Okrem toho je mastoid spojený s optickými tuberkulózami a retikulárnou tvorbou stredného mozgu. Pes má spárované telo.
vývoj medzimozgu
Diencefalon je položený z malého počtu buniek v stene obrovskej tretej srdcovej komory mozgu, iba u suchozemských zvierat. A to najmä u cicavcov dosahuje značnú veľkosť.
1. Epitalamus tvorí dobre vyjadrená embryonálna doska pneumatiky u všetkých dospelých cicavcov. Šišinka je základom tretieho nepárového parietálneho oka. Iba niektoré vodné živočíchy a plazy majú pod kožou močový mechúr. U cicavcov sa epifýza stáva endokrinnou žľazou, ktorej funkcia ešte stále nie je dostatočne objasnená.
2. Zvláštna a vysoko vyvinutá embryonálna doštička dna tvorí subtalamskú časť diencefalonu. Pochádza z epichordálneho mozgu, to znamená, že je spojený so stredným mozgom.
Hypofýza pochádza z troch rôznych zdrojov. Hrtanová ektoderma sa zmení na rozvetvenú žľazu. Lúmen žľazy následne zmizne, zostávajú však vlákna glandulárnych buniek rôznych štruktúr, obklopené veľkým počtom ciev. U suchozemských zvierat sa zo steny lievika objaví nervová časť hypofýzy z nervových a gliových buniek a nervových vlákien. Izoluje sa stredná časť hypofýzy z epitelových buniek. Hypofýza ako celok vylučuje viac ako tucet rôznych hormónov, ktoré z časti žľazy vstupujú do krvného riečišťa a od nervových a stredných častí do mozgovomiechového moku. Hypofýza je v interakcii s autonómnymi centrami zabudovanými v stenách tretej srdcovej komory.
3. Embryonálna bočná platňa tvorí vizuálny borax-talamus - a očné vezikuly. Štruktúra talamu zahŕňa jadrá optických tuberkulóz. Slúžia:
a) prostredné strediská všetkých ciest, ktoré smerujú do mozgovej kôry a vykonávajú rôzne impulzy - čuchové, citlivosť kože a svalov, chuť a u suchozemských zvierat navyše vizuálne a statoakustické;
b) stredný stred všetkých ciest, ktoré prechádzajú od kôry plášťa do rôznych častí mozgu.
To vysvetľuje, prečo sa vizuálne hľuzy začínajú tvoriť s čuchovým plášťom a dosahujú maximálny vývoj u cicavcov v súvislosti s tvorbou sekundárneho plášťa. Silný vývoj optických hľúz je tiež spôsobený pohybom vizuálnych centier od stredného mozgu k sekundárnemu plášťu v dôsledku spojenia s mozočkom. V dôsledku obohatenia talamu o stredné stredy rastie stredná hmota optických tuberkúl, ktorá sa po zavedení do dutiny tretej srdcovej komory v mozgu zmení na kruhový kanálik. V šedej hmote steny tretej mozgovej komory sú početné vyššie subkortikálne autonómne centrá.
Stredná mozog-Mesencephalon
Skladá sa z: štvornásobnej platne, čiapky nôh alebo kapoty z nôh veľkého mozgu. Dutina stredného mozgu sa zmenila na mozgový akvadukt aqueductus cerebri - spája tretiu a štvrtú mozgovú komoru. Centrálna šedá meduľa pneumatiky je zabudovaná v stenách prívodu vody.
1. doštička so štvorcovým tektom - predstavuje dorzálnu časť stredného mozgu, leží kaudálne z vizuálnych hľúz a nazálne z mozočku. Doska pozostáva z párových nazálnych a kaudálnych kopcov - colliculi nasalis et caudalis. Kopce sú oddelené priečnymi a strednými drážkami. Z povrchu je štvornásobná doska potiahnutá látkou z bieleho mozgu, pod ktorou sa nachádza šedá látka z mozgu, v nosných kopcoch je to subkortikálne centrum rôznych aferentných dráh vrátane vizuálnych a v kaudálnych, subkortikálnych zvukových a rovnovážnych centrách.
Všeobecne platí, že štvornásobná platňa je koordinačným centrom mnohých impulzov: čuchový, vizuálny, rovnovážny, zvukový, všeobecná citlivosť a impulzy z mozgovej kôry. Motorické impulzy sa prenášajú na červené jadro, miechu, ako aj na hlavné svaly, na mozoček a na mostík.
2 - v strede stredného mozgu medzi štvoritou platňou a nohami veľkého mozgu poťah nôh alebo cap-tegmentum pedunculi , V nej ležia párové jadrá šedej drene, v rovine predných kopcov:
a) centrum kormidla červeného jadra a jadra miechy;
b) jadro jadra okulomotorického nervu n. oculomotorius.
c) parasympatické jadrá Jakuboviča.
Kaudálne umiestnené:
d) jadro blokového nervového jadra n. trochlealis;
e) časť jadra piateho nervu.
Retikulárna formácia prechádza cez celú čiapočku od medulla oblongata k medziproduktu a vytvára cesty od miechy a mozočka do štvorkolky, do vizuálnych hľúz a od nich do miechy.
3.leg cerebrum-pedunculi cerebri - sa objavia na bazálnom povrchu mozgu vo forme dvoch hrubých hrebeňov medzi optickými traktmi a mozgovým mostíkom. Sú rozdelené drážkou medzi nohami. Z nôh vystupujú 3 páry lebečných nervov - okulomotorický nerv. Nohy sú tvorené efferentnými cestami, ktoré spájajú plášťovú kôru s kosoštvorcom a miechou. Nohy sú preto vyvinutejšie u tých zvierat, v ktorých je mozgová kôra silnejšia. U ľudí sú najrozvinutejšie.
Vývoj stredného mozgu
Stredná mozog u nižších zvierat, rovnako ako u embryí vyšších zvierat, dosahuje veľmi významnú veľkosť. Šedá medulla embryonálnej laterálnej laminy stredného mozgového mechúra, ktorá rastie, tvorí oblúk stredného mozgu, z ktorého sa u spodných zvierat vyskytuje dolná kolónia alebo vizuálne laloky. Spočiatku bolo dicollis najvyšším koordinačným centrom, pretože v ňom končia nielen optické nervy, ale aj aferentné dráhy od striatum, kosoštvorca a miechy. Plazy však už majú nejaké vlákna, zatiaľ čo cicavce majú takmer všetky vlákna zrakový nerv pohybujú sa cez vizuálne tuberkulózy diencephalonu do kôry sekundárneho plášťa. Preto vizuálne laloky zaostávajú v raste a sekundárny plášť a teda aj vizuálne hľuzy rastú čoraz viac.
U suchozemských zvierat v oblúku stredného mozgu sú položené rovnovážne zvukové centrá, spočiatku vo forme mikroskopických útvarov a neskôr makroskopických útvarov. Iba u cicavcov sa namiesto dichotómie objaví kvadrupól. U zvierat s dobrým sluchom prevládajú vrchné zvukovody.
Z embryonálnej hlavnej doštičky vo ventrálnej stene stredného mozgového mechúra sa vytvorí viečko. V dôsledku tvorby oblúka a viečka sa dutina stredného mozgu zmení na mozgový akvadukt. Čiapka pozostáva z jadier 3 a 4 párov lebečných nervov a špeciálnych motorových jadier čiapky. Tieto zahŕňajú červené jadro spájajúce mozoček s miechou a intersticiálne jadro spájajúce ganglion frenula s čuchovým mozgom.
U cicavcov je čiapka v spojení s vývojom sekundárneho plášťa ventrálne prerušená vrstvou bielej mozgovej hmoty z ciest vedúcich od kôry k kosoštvorcom a mieche. Tieto dráhy tvoria nohy mozgu, ktorých sila zodpovedá stupňu rozvoja mozgovej kôry.
Zadný mozog-Rhombencephalon
Rozdelené na drôty oblongata a hindbrain. Zadný mozog sa skladá z mozgu a mozgového mosta. Medzi mozočkom a podlhovastým mostom je štvrtá mozgová komora.
A) Medulla oblongata-medulla oblongata
Pokračuje kaudálne bez zreteľného ohraničenia miechy. Na bazálnom povrchu je zreteľne viditeľný stredný ventrálny sulcus –fissura mediana ventralis. Na oboch jeho stranách prechádzajú bočné drážky - sulcus paramedianus - kaudálne tečú do strednej ventrálnej drážky. Medzi týmito tromi brázami sú dva úzke hrebene - pyramídy - pyramídy, v ktorých prechádzajú pyramidálne vodivé zväzky z mozgovej kôry do miechy. Pretože vodivé zväzky pokračujú do bočných kordov a miechy a súčasne prechádzajú sprava doľava a naopak, vytvára sa pyramídovo-decussatio-pyramidový kríž. Neskôr z pyramídy za mostom prichádzajú 4 páry lebečných nervov - únosový nerv. Blízko priesečníka pyramíd a 12 párov sa od nej laterálne odchýlia - hyoidný nerv, ale ďalšie dva nervy, jeden po druhom, z hyoidného nervu laterálne: 10 para-vagus nervu, viac nazálne-9 para-leskofaryngeálny nerv.
Medulla oblongata je zoskupená:
a) v samostatných senzorických a motorických jadrách, z ktorých pochádza 5, 6,7,9,10 a 12 párov lebečných nervov;
b) do stredných jadier lúčov Gaulla a Burdach;
c) v jadre je 8 párov as nimi spojené nosové a kaudálne olivy-7.
Medzi jadrami leží sieťovina - formatio reticularis - z prepletených nervových vlákien a nervových buniek medzi nimi, ktoré z drene oblongata prechádzajú do čiapky stredného mozgu a do diencefalonu. Vykonáva predovšetkým asociačnú a koordinačnú funkciu medzi rôznymi jadrami kosoštvorca a stredného mozgu a je centrom dýchania a kardiovaskulárneho systému.
Biela drieň medulla oblongata pozostáva z veľkého počtu zväzkov ciest vedúcich od miechy k rôznym častiam mozgu a naopak.
B) Cerebellum-cerebellum
Má takmer guľový tvar, s dvoma drážkami je rozdelený na strednú časť - červa a dve bočné laloky. Šedá medulla tvorí mozgovú kôru-kôra mozgu - a jednotlivé jadrá ležiace v centrálne umiestnenej bielej hmote mozgu. Funkcia - koordinácia pohybov.
Povrch kôry červieho vermisu je rezaný priečnymi drážkami a trhlinami. Červ je s dvoma hlavnými trhlinami ohraničený na predné, stredné a zadné laloky - lobus rostralis, caudalis et medialis. Každá z nich je spojená s nohami mozočku - predným, stredným a zadným, skladajúcimi sa z chodníkov. Predné a zadné konce závitovky sú ohnuté ventrálne a k sebe, medzi nimi zostáva malá medzera - horná časť stanu-fastigium. Látka bieleho mozgu červa-7 na sagitálnej sekcii pripomína vetvu thuja, preto sa nazýva strom života-altánok vitae. V bielej mozgovej látke červa sa nachádza stanové jadro, nucleus fastigii, ktoré je subkortikálnym centrom analyzátora rovnováhy.
Pri medulle oblongata sa červ spája cez kaudálny mozgový šalát-velum medullare caudale a so štvorkolkou cez nosový mozgový šalát-velum medullare nasale.
Mozgová hemisféra - hemisphaerum cerebelli - ako červ sa skladá z početných lalokov, z ktorých jedna je drvená vločka, vo forme malého prívesku červa, ktorý leží za bočným pediklom. V bielej drene mozgových hemisfér sú jadrá dentátového jadra, ktoré slúžia ako prenosové centrá motorických impulzov.
Mozoček sa pripája k medulle oblongata. S mozgovým mostíkom - bočnými nohami a stredným mozgom - nosnými nohami.
Na dorzálnom povrchu medulla oblongata vyčnievajú chvostové nohy cerebellum-pedunculus cerebellaris medullum oblongatum - vo forme dvoch hrebeňov. Cesty nimi prechádzajú:
a) z miechy - zväzky Flexig, Gaulle a Burdach s ich jadierami;
b) z jadier sluchového nervu;
c) z kaudálnych olív;
d) z jadier 5,10 a 12 párov lebečných nervov.
Nosné nohy mozočka-brachia conum ctivum cerebelli idú pod zadné kopce štvornásobku do nôh mozgu. Nosné nohy ležia na chrbtovom povrchu drene oblongata. Cesty nimi prechádzajú:
a) z miechy do zväzku červov Hovers;
b) z dentálnych jadier mozgovej hemisféry do červeného jadra;
c) v jadre vizuálnych hľúz.
Vnútorné dráhy mozgu sú predstavované vláknami medzi mozgovou kôrou a jej jadrami a asociatívnymi vláknami spájajúcimi oddelené gyrusy v sagitálnej rovine.
C) Mozgové mostíky z mozgu
Leží na prednom konci medulla oblongata, na hranici so strednou mozgovou bruškou, v tvare priečneho hrebeňa, ktorý sa na svojich koncoch ohýba chrbtovo k mozočku a tvorí bočné končatiny cerebellum-brachium pontis. Most a nohy pozostávajú z ciest spájajúcich jadrá jadra mosta pontis s jadierami mozgu. Na jadrách mosta začínajú cesty od konca mozgovej kôry a cesty k mozgovým hemisférom. Bočnými úsekmi mosta vystupuje 5 párov - trigeminálny nerv - Najmasívnejší nerv zo všetkých lebečných nervov. Má dva korene: ventrálny - nazálny motor a dorso - kaudálny senzorický systém. Na poslednom je veľký lunárny ganglion. Z mosta, tiež v priečnom smere, leží lichobežníkové teleso, corpus trapezoideum, v tvare úzkeho a nízkeho hrebeňa. Je tvorená dráhami vedúcimi z jadier sluchového nervu. 8 para-rovnovážno-sluchový nerv a 7 para-faciálny nerv vystupujú cez bočné časti lichobežníkového tela.
D) Štvrtý cerebrálny komorový-ventrikulárny kvartus
Umiestnené medzi mozoček a drieň oblongata. Oblúk je podávaný červami a mozgovými plachtami a spodná časť medulla oblongata.
Spodok štvrtej komory sa nazýva kosoštvorec fossa-fossa rhomboidea. Medián a dve bočné drážky v dolnej časti fossy sú párové stredné vyvýšeniny, eminentia medialis, na ktorých stojí maska \u200b\u200btváre - colliculi facialis - proti laterálnemu pedikulu mozočka. V oblasti tváre mohyly ležia jadrá abdukčných a tvárových nervov. Na kaudálnom konci strednej výšky je pole hypoglossu hyoidnej nervovej oblasti s jadrom rovnakého mena.
Jadro 9 a 10 párov nervov vyčnieva laterálne z hyoidného nervu. Tvoria sivé krídlo ala cinerea. Oblasť zadného konca sivých krídel je známa ako pen-calamus scriptorius.
Priamo za postrannými nohami mozočka a stredne od nich sa vestibulárne polia oblasti objavujú vo forme malých výšok. Obsahujú vestibulárne a kochleárne jadrá 8 párov nervov. Kochleárne jadra ležia laterálne.
Vývoj kosoštvorcov
Medulla oblongata sa vyvíja proliferáciou laterálnych a hlavných doštičiek embryonálnej mozgovej trubice. V dôsledku rozšírenia bočných dosiek formovacím mozkom boli stredy motorických centier medully oblongata mediálne posunuté a zmyslové senzory boli laterálne posunuté.
To isté je spôsobené strednou polohou na driekovej oblongate motorických nervov a laterálne citlivými, ako aj štrukturálnymi črtami spodnej časti štvrtej srdcovej komory mozgu, v hrúbke ktorej citlivé jadrá ležia bočne od motora.
Štruktúra medulla oblongata všeobecne u zvierat je makroskopicky dosť jednotná, čo sa vysvetľuje jej jednotnou funkciou - stredy žiabrových prístrojov a ich derivátov, črevá, stredy dotykových orgánov, kĺbovo-svalové pocity, sluch a rovnováha. Vzhľadom na to, že cesty spájajúce mozog s miechou prechádzajú cez drieň oblongata, je jej šedá drieň rozdelená do samostatných jadier, na rozdiel od kompaktnej šedej drene miechy.
Pri medulle oblongata si zasluhuje osobitnú pozornosť motorické jadro formovania čiapky alebo oka, ktoré sa prvýkrát vyskytuje u rýb a je najstarším združením a motorickým centrom. U suchozemských zvierat sa nosné a kaudálne olivy vytvárajú ako asociačné centrá kvôli motorickému jadru čiapky. Nosové olivy sa objavujú iba u suchozemských zvierat, počnúc obojživelníkmi. Slúžia ako medziľahlé centrum na ceste od kochleárneho nervu k optickým tuberkulám. Zväzok týchto vlákien tvorí lichobežníkové teleso. U vtákov a cicavcov sa kaudálne olivy tvoria ešte neskôr. Sú spojené s jadierami chrbtových kordov, s vizuálnymi tuberkulózami, s mozgom a miechou. Vo svojich vzťahoch sú kaudálne olivy najužšie spojené s funkciou udržiavania rovnováhy.
Mozoček sa vyvíja v súvislosti s jeho funkciou udržiavania rovnováhy tela a udržiavania svalového tonusu. Preto je najvýraznejší u zvierat, ktoré rýchlo plávajú, behajú, skákajú, lietajú a sú slabšie u zvierat, ktoré sa pohybujú pomaly. V primitívnej forme predstavuje mozoček nepárový tanier a biela alebo sivá medulla. Mozoček je položený uprostred embryonálnej platne krytu kosoštvorca a predné a zadné mozgové plachty sú tvorené z prednej a zadnej časti druhej. Cerebelárna doska, ktorá rastie spredu dozadu, sa oblúkovite ohýba v dorzálnom smere. V dôsledku objavenia sa pozdĺžnych drážok je izolovaná stredná časť doštičky, telo mozgu a párové bočné časti, uši mozočka. Telo mozgu a uši sú paleocerebellum. Telo mozočka u suchozemských zvierat je rozdelené priečnymi drážkami na predné, stredné a zadné laloky, na ktorých sa objavujú ďalšie priečne drážky. Predný lalok je spojený so svalmi hlavy a stredný a zadný lalok sú spojené so svalmi trupu a končatín.
U cicavcov prevažuje priemerný podiel. Pozdĺžne drážky na ňom oddeľujú strednú nepárovú časť - červ - od párových bočných lalokov alebo od mozgových hemisfér. Mozgové hemisféry sú neocerebellum. Červ obsahuje stredy koordinovaných, synchrónnych pohybov kmeňa a končatín. Mozgové hemisféry sú najvýraznejšie u vyšších cicavcov, ktoré majú vo väčšej alebo menšej miere schopnosť oddeliť pohyby končatín. Zlepšenie tejto schopnosti zase závisí od sily mozgovej kôry ako najvyššieho centra nervovej aktivity a od výskytu mozgu s mozgovou kôrou cez jeho bočné nohy a mostík.
Mozgové hemisféry a most sa nachádzajú iba u cicavcov a dosahujú maximálny vývoj u primátov. Homogénna funkcia mozočka u zvierat sa vysvetľuje pomerne uniformnou histologická štruktúra jeho kôra, v ktorej je rozlíšená povrchová molekulárna vrstva, hlboká granulárna a stredná vrstva veľkých Purkínových buniek.
Uši primitívneho mozočka u vodných živočíchov súvisia s rovnovážnymi orgánmi, tj s orgánmi bočnej línie as svalmi chvosta. S redukciou týchto orgánov u suchozemských zvierat uši klesajú. U cicavcov sa z nich zachovávajú kúsky, ktoré sa spájajú so zadným lalokom červa.
Ongenéza červa je tiež určená jeho funkciou, ktorá je zvlášť viditeľná v ľudskom mozgu. U novonarodeného dieťaťa je stále veľmi malý a predstavuje iba 1 / 16-1 / 18 hmotnosti veľkého mozgu. Ale tieto pomery sa menia s rastúcim dieťaťom a zvyšuje sa jeho mobilita, u dospelých sú pomery už rôzne - 1 / 8-1 / 9 hmotnosti mozgu.
Zoznam použitej literatúry
1) A.I. Akayevsky "Anatomy of Pets"
2) I.V. Khrustaleva "Anatomy of Pets"
3) L.V. Antipova, V.S. Slobodyanik, S. M. Suleymanov „Anatómia a histológia hospodárskych zvierat“
4) A.F. "Anatómia domácich zvierat" Rimov
5) A.F. Kuznetsova „Sprievodca veterinárnym lekárom“