Opíšte hlavné typy svalového tkaniva. Histologická štruktúra svalového tkaniva.
Svalové tkanivo sa nazýva tkanivo, ktoré sa líši v štruktúre a pôvode, ale podobné schopnosti prejavovať kontrakcie. Poskytujú pohyb v celom organizme ako celku alebo jeho častiach (napríklad kostrové svaly) a pohyb orgánov vo vnútri orgánu (napríklad srdce, jazyk, črevá).
Bunky mnohých tkanív majú vlastnosť zmeny tvaru, ale táto schopnosť sa stáva hlavnou funkciou svalového tkaniva.
Všeobecná charakteristika a klasifikácia
Hlavnými morfologickými znakmi prvkov svalového tkaniva sú pretiahnutý tvar, prítomnosť pozdĺžne umiestnených myofibríl a myofilamentov - špeciálne organely, ktoré zabezpečujú kontraktilitu, lokalizáciu mitochondrií v blízkosti kontraktilných prvkov, prítomnosť inklúzií glykogénu, lipidov a myoglobínu.
Hladké svalové tkanivo.
Toto tkanivo je tvorené mezenchýmom. Bunky hladkého svalstva - myocyty pretiahnuté špicatými koncami fusiform, pokryté bunkovou membránou. Myocyt je mononukleárna bunka. Jadrové jadro zaujíma centrálnu polohu v bunke. Obsahuje atomizovaný heterochromatín a jeden alebo dva jasne viditeľné jadrá. Bunkové centrum (cytocenter) je umiestnené blízko jedného z povrchov jadra. Zostávajúce organely sa koncentrujú na póloch jadra v tvare tyčinky. Dobre vyvinutá mitochondria, Golgiho komplex, hladký EPS; ribozómy, granulovaný EPS sú mierne exprimované. Cytoplazma myocytov obsahuje glykogénové granule - energetickú rezervu bunky.
Myocyty tesne priliehajú k sebe a tvoria vrstvy a skupiny, oddelené voľným netvoreným spojivovým tkanivom. Myofibrily sa nachádzajú v cytoplazme, idú pozdĺž obvodu bunky pozdĺž jej osi. Pozostávajú z tenkých nití a sú kontraktívnym prvkom svalu.
Vonku je svalové vlákno pokryté membránou - sarkolemou, ktorá sa skladá z vnútornej a vonkajšej vrstvy. Vnútornou vrstvou je plazmolém, ktorý je podobný membráne iných tkanivových buniek. Vonkajšia spojivová tkanivová vrstva sa skladá z bazálnej membrány a susedných vláknitých štruktúr. Plazmoléma tvorí systém úzkych tubulov, ktoré prenikajú do svalových vlákien.
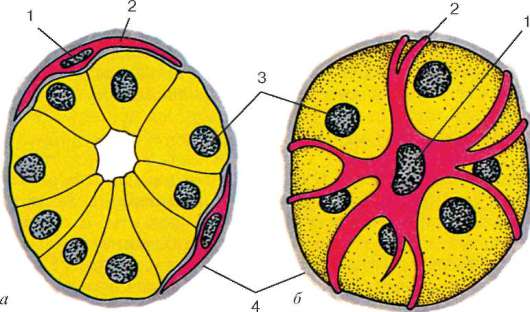
Spojivové tkanivové vlákna umiestnené mimo bazálnej membrány svalového vlákna tvoria endomýzium, ktoré je bohaté na krvné cievy a nervy. Endomysium sa kombinuje s perimisiom, membránou pokrývajúcou skupinu svalových vlákien. Perimisium niekoľkých svalových zväzkov je spojené s epimisiom - vonkajšou membránou spojivového tkaniva, ktorá kombinuje niekoľko týchto zväzkov do svalu - orgánu charakterizovaného špecifickou štruktúrou a funkciou.
Hladké svalové tkanivo sa nachádza v stenách žalúdka, čriev, maternice, mechúr, močovodov, priedušiek, ako aj v stredných a veľkých krvných cievach. Aktivita hladkého svalstva je regulovaná pomocou ANS. Svalové kontrakcie neposlúchajú vôľu osoby, a preto sa hladké svalové tkanivo nazýva nedobrovoľné svalstvo.
Pružné svalové tkanivo.
Kostrové svalové tkanivo je sťahovacie tkanivo trupu, hlavy, končatín, hltanu, hrtanu, hornej polovice pažeráka, jazyka a žuvacích svalov.
Tvorí sa z myotómov somitov mezodermu. Štrukturálna jednotka je pruhované svalové vlákno. Má valcovité telo, je pokryté škrupinou - sarkolem a cytoplazma sa nazýva - sarkoplazma, v ktorej je veľa jadier a myofibríl. Každý myofibril pozostáva z diskov s rôznym chemickým zložením. Myofibrily sú kontraktívnym aparátom svalových vlákien.
Celé kostrové svaly sú vytvorené z priečne pruhovaného svalového tkaniva. Svaly sú ľubovoľné, pretože k jeho kontrakcii dochádza pod vplyvom mozgovej kôry.
Svalové tkanivo srdca.
Tento typ svalového tkaniva tvorí strednú membránu srdca - myokard sa povahou redukcie vzťahuje na nedobrovoľné, pretože nie je kontrolovaná vôľou zvieraťa. Vyvíja sa z viscerálneho listu splanchnatómu - myoepikardiálnej laminy.
Srdcové svalové tkanivo pozostáva zo svalových buniek - kardiomyocytov (srdcové myocyty). Myocyty, ktoré sa navzájom spájajú na svojich koncoch pozdĺž dlhej osi buniek, tvoria štruktúru podobnú svalovému vláknu.
Existujú dva typy buniek. : typické kontraktilné bunky a atypické srdcové myocytytvoriaci vodivý systém srdca.
Typické svalové bunky plniť zmluvnú funkciu; majú pravouhlý tvar, 1-2 jadrá sú v strede, myofibrily sú umiestnené na periférii. Medzi susednými myocytmi sú vložené disky. S ich pomocou sa myocyty zhromažďujú vo svalových vláknach oddelených tenkým vláknitým spojivovým tkanivom. Medzi susednými svalovými vláknami sú spojovacie vlákna, ktoré poskytujú kontrakciu myokardu ako celku.
Vodivý systém srdca je tvorený svalovými vláknami, ktoré sa skladajú z atypické svalové bunky, Sú väčšie ako kontraktilné, bohatšie na sarkoplazmu, ale chudobnejšie ako myofibrily, ktoré sa často pretínajú. Jadrá sú väčšie a nie vždy v strede. Vlákna vodivého systému sú obklopené hustým plexom nervových vlákien.
Svalové tkanivá (lat. Textus muscularis) sa nazývajú tkanivá, ktoré sa líšia štruktúrou a pôvodom, ale majú podobnú schopnosť výrazných kontrakcií. Svalové tkanivo je tvorené predĺženými bunkami, ktoré spôsobujú podráždenie nervový systém a reagovať so znížením. Poskytujú pohyb v celom tele tela, pohyb orgánov vo vnútri tela (srdce, jazyk, črevá atď.) A sú tvorené svalovými vláknami. Bunky mnohých tkanív majú vlastnosť zmeny tvaru, ale vo svalových tkanivách sa táto schopnosť stáva hlavnou funkciou.
Hlavné morfologické znaky prvkov svalového tkaniva: predĺžený tvar, prítomnosť pozdĺžne umiestnených myofibríl a myofilamentov - špeciálne organely, ktoré zabezpečujú kontraktilitu, lokalizáciu mitochondrií v blízkosti kontraktilných prvkov, prítomnosť inklúzií glykogénu, lipidov a myoglobínu.
Špeciálne kontraktilné organely, myofily alebo myofibrily poskytujú kontrakciu, ku ktorej dochádza pri interakcii dvoch hlavných fibrilárnych proteínov - aktínu a myozínu, s povinnou účasťou iónov vápnika. Mitochondrie dodávajú týmto procesom energiu. Zásobu energetických zdrojov tvoria glykogén a lipidy. Myoglobín je proteín, ktorý zaisťuje viazanie kyslíka a vytváranie jeho rezervy v čase kontrakcie svalov, keď sú krvné cievy stlačené (prísun kyslíka prudko klesá).
Vlastnosti svalového tkaniva:
- vzrušivosť
- vodivosť
- vtažitelnost
- labilita
Druhy svalového tkaniva:
1. Hladké svalové tkanivo
Tkanivo hladkého svalstva sa skladá z jednojadrových buniek - vretenovitých myocytov s dĺžkou 20 - 500 mikrónov. Ich cytoplazma vo svetelnom mikroskope vyzerá jednotne, bez priečneho ryhovania. Toto svalové tkanivo má špeciálne vlastnosti: pomaly sa sťahuje a uvoľňuje, má automatickosť, je nedobrovoľné (to znamená, že jeho činnosť nie je kontrolovaná vôľou osoby). Zahrnuté v stenách vnútorné orgány: krvné a lymfatické cievy, močové cesty, tráviaci trakt (kontrakcia stien žalúdka a čriev).
2. Pružené tkanivo kostrového svalstva
Skladá sa z myocytov, ktoré majú veľkú dĺžku (až niekoľko centimetrov) a priemer 50 až 100 mikrónov; tieto bunky sú viacjadrové, obsahujú až 100 alebo viac jadier; v svetelnom mikroskope vyzerá cytoplazma ako striedanie tmavých a svetlých pruhov. Vlastnosti tohto svalového tkaniva sú vysoká miera kontrakcie, relaxácie a náhodnosti (to znamená, že jeho činnosť je kontrolovaná vôľou osoby). Toto svalové tkanivo je súčasťou kostrových svalov, rovnako ako steny hltanu, horná časť pažeráka, tvorí jazyk, okulomotorické svaly. Vlákna dlhé 10 až 12 cm.
3. Pružené tkanivo srdcového svalu
Skladá sa z 1 alebo 2 jadrových kardiomyocytov, ktoré majú priečne pruhovanie cytoplazmy (pozdĺž periférie cytolémie). Kardiomyocyty sú rozvetvené a vytvárajú vzájomne prepojené spojenia - vkladacie disky, v ktorých sa kombinuje ich cytoplazma. Existuje aj ďalší medzibunkový kontakt - anostamózy (inhibícia cytolémie jednej bunky do cytolémie druhej). Tento typ svalového tkaniva tvorí myokard srdca. Vyvíja sa z myoepikardiálnej doštičky (viscerálny list splanchnotómu embrya na krku). Osobitnou vlastnosťou tohto tkaniva je automatizácia - schopnosť rytmicky sa sťahovať a relaxovať pod vplyvom excitácie, ktorá sa vyskytuje v samotných bunkách (typické kardiomyocyty). Toto tkanivo je nedobrovoľné (atypické kardiomyocyty). Existuje tretí typ kardiomyocytov - sekrečné kardiomyocyty (nemajú vlákna), ktoré syntetizujú hormón troponín, ktorý znižuje krvný tlak a rozširuje steny krvných ciev.
Svalová srdcová membrána (myokard) je tvorená bunkami svalovej dutiny, ktoré sú pomocou zavádzacích diskov spojené do svalových komplexov alebo srdcových svalových vlákien. Svalové tkanivo má schopnosť regenerácie, je chránené spojivovým tkanivom a vytvára jazvu.
Odovzdať svoju dobrú prácu do vedomostnej základne je ľahké. Použite nasledujúci formulár
Študenti, absolventi vysokých škôl, mladí vedci, ktorí vo svojich štúdiách a práci využívajú vedomostnú základňu, vám budú veľmi vďační.
Publikované dňa http://www.allbest.ru/
Svalové tkanivo kombinuje schopnosť redukcie.
Štrukturálne vlastnosti: kontraktilný prístroj, ktorý zaberá významnú časť v cytoplazme štrukturálnych prvkov svalového tkaniva a pozostáva z aktínových a myozínových vlákien, ktoré tvoria organely na špeciálne účely - myofibrils .
Svalové tkanivoide o skupinu tkanív rôzneho pôvodu a štruktúry spojenú na základe spoločného znaku - výraznej kontraktilnej schopnosti, vďaka ktorej môžu vykonávať svoju hlavnú funkciu - pohybovať telom alebo jeho časťami v priestore.
Najdôležitejšie vlastnosti svalového tkaniva.Štrukturálne prvky svalového tkaniva (bunky, vlákna) majú podlhovastý tvar a sú schopné sťahovania kvôli silnému vývoju kontraktilného aparátu. Ten sa vyznačuje vysoko usporiadaným usporiadaním aktínua myozínové myofily,vytváranie optimálnych podmienok pre ich interakciu. Toto je dosiahnuté spojením kontraktilných štruktúr so špeciálnymi prvkami cytoskeletu a plazmolému (Sarkolemou)predvádzanie referenčná funkcia, V časti svalového tkaniva tvoria myofily organely osobitného významu - myofibrils.Na svalovú kontrakciu je potrebné značné množstvo energie, preto v štrukturálnych prvkoch svalových tkanív existuje veľké množstvo mitochondrií a trofických inklúzií (lipidové kvapky, glykogénové granule) obsahujúce substráty - zdroje energie. Pretože svalová kontrakcia nastáva za účasti iónov vápnika, štruktúry, ktoré vykonávajú jej akumuláciu a sekréciu, sú dobre vyvinuté vo svalových bunkách a vláknach - agranulárnom endoplazmatickom retikule. (sarkoplazmatické retikulum), jaskyňa.
Klasifikácia svalového tkanivana základe znakov ich (a) štruktúry a funkcie (morphofunkčná klasifikácia)a b) pôvod (histogénna klasifikácia).
Morphofunkčná klasifikácia svalového tkaniva zdôrazňuje priečne pruhované (priečne pruhované) svalové tkanivoa tkanivo hladkého svalstva.Prúžkované svalové tkanivo je tvorené štruktúrnymi prvkami (bunky, vlákna), ktoré majú priečne pruhovanie v dôsledku špeciálneho usporiadaného vzájomného umiestnenia aktínu a myozínových myofilov v nich. Prúžkové svalové tkanivá zahŕňajú skeletováa tkanivo srdcového svalu.Hladké svalové tkanivo pozostáva z buniek, ktoré nemajú priečne pruhovanie. Najbežnejším typom tohto tkaniva je tkanivo hladkého svalstva, ktoré je súčasťou steny rôznych orgánov (priedušiek, žalúdok, črevá, maternica, vajcovod, močovod, močový mechúr a krvné cievy).
Histogenetická klasifikácia svalového tkaniva rozlišuje tri hlavné typy svalového tkaniva: somatická(kostrové svalové tkanivo) coelomic(srdcové svalové tkanivo) a mezenchymálnych(tkanivo vnútorných orgánov hladkého svalstva), ako aj ďalšie dva: myoepiteliálne bunky(upravené epitelové kontraktilné bunky v koncových rezoch a malé vylučovacie kanáliky niektorých žliaz) a minerálne prvky(kontraktívne bunky nervového pôvodu v dúhovke oka).
Kostné pruhované (pruhované) svalové tkanivovo svojej hmotnosti presahuje akékoľvek iné tkanivo tela a je najbežnejším svalovým tkanivom ľudského tela. Poskytuje pohyb tela a jeho častí v priestore a udržuje držanie tela (súčasť pohybového aparátu), vytvára okulomotorické svaly, svaly steny ústnej dutiny, jazyk, hltan a hrtan. Neskeletálne viscerálne pruhované svalové tkanivo, ktoré sa nachádza v hornej tretine pažeráka, je súčasťou vonkajších análnych a uretrálnych zvieračov.
svalový kostrový myocyt
Svalové tkanivo pruhovaného svalstva sa vyvíja v embryonálnom období od myotomessomiti, ktorí vedú k aktívnemu rozdeleniu myoblasty- bunky, ktoré sú usporiadané v reťazcoch a spájajú sa navzájom v oblasti koncov s formáciou svalové kanáliky (myotubuly)premenil sa na svalové vlákna.Takéto štruktúry tvorené jednou obrovskou cytoplazmou a početnými jadrami sa v ruskej literatúre tradične označujú ako symplasts(v tomto prípade - myosymplasts)v akceptovanej medzinárodnej terminológii však tento pojem chýba. Niektoré myoblasty sa nezlučujú s ostatnými, nachádzajúcimi sa na povrchu vlákien a spôsobujúcimi vznik miosatellitotsitam- malé bunky, ktoré sú cambiálnymi prvkami tkaniva kostrového svalstva. Kostrové svalové tkanivo tvorené vo zväzkoch pruhované svalové vlákna, ktoré sú jej štrukturálne a funkčné jednotky.
Svalové vlákna tkanivo kostrového svalstva sú valcovité útvary rôznej dĺžky (od milimetrov do 10 - 30 cm). Ich priemer sa tiež veľmi líši v závislosti od príslušnosti k určitému svalu a typu, funkčného stavu, stupňa funkčnej záťaže, stavu výživy a ďalších faktorov. Vo svaloch tvoria svalové vlákna zväzky, v ktorých ležia rovnobežne a pri vzájomnom zdeformovaní často získajú nepravidelný mnohotvárny tvar, ktorý je zvlášť zreteľný v priečnych rezoch. Medzi svalovými vláknami sú tenké vrstvy sypkých vlákien spojivové tkanivoprenášajúce cievy a nervy - endomysium.Priečne pruhovanie vlákien kostrových svalov v dôsledku striedania tmy anizotropné disky (pásy A)a svetlé izotropné disky (pásyI). Každý izotropický disk je rozrezaný na polovicu tenkou tmavou farbou linka Z - telefragm, Jadrá svalového vlákna - relatívne ľahké, s 1 - 2 jadierami, diploidnými, oválnymi, sploštenými - ležia na jeho obvode pod sarkolemou a sú umiestnené pozdĺž vlákna. Mimo sarkolemmu je pokrytý hustou vrstvou bazálna membránado ktorých sú tkané retikulové vlákna.
Myosatelitné bunky (myosatelitné bunky) - malé sploštené bunky umiestnené v plytkých dojmoch sarkolemmy svalového vlákna a pokryté spoločnou bazálnou membránou (pozri obrázok 88). Jadro myosatelitu je husté, pomerne veľké, organely sú malé a málo. Tieto bunky sa aktivujú, keď sú poškodené svalové vlákna a poskytujú ich regeneratívnu regeneráciu. Zlúčením so zvyškom vlákna so zvýšenou záťažou sú myosatelitné bunky zapojené do jeho hypertrofie.
myofibrils z kontraktilného aparátu svalového vlákna, sú umiestnené v sarkoplazme pozdĺž svojej dĺžky, zaberajú strednú časť a sú zreteľne odhalené na priečnych úsekoch vlákien vo forme malých bodiek.
Myofibrily majú svoje vlastné priečne pruhovanie a vo svalovom vlákne sú usporiadané tak, že sa navzájom zhodujú izotropné a anizotropné disky rôznych myofibríl, čo spôsobuje priečne pruhovanie celého vlákna. Každý myofibril je tvorený tisíckami opakujúcich sa postupne prepojených štruktúr - sarkomérov.
inokomma (Myomere)je štruktúrna a funkčná jednotka myofibríl a predstavuje jej časť umiestnenú medzi dvoma ťahače (riadky Z).Obsahuje anizotropný disk a dve polovice izotropných diskov - jednu polovicu na každej strane. Sarcomere tvoril objednaný systém hrubý (myozín)a tenké (aktínové) vlákna.Hrubé myofily sú spojené mezofragmus (čiara M)a koncentrovaný v anizotropnom disku,
a tenké myofily spojené telogramy (riadky Z),tvoria izotropné disky a čiastočne prenikajú do anizotropného disku medzi hrubými vláknami až do svetla pás Hv strede anizotropného disku.
Vo svaloch, rovnako ako v iných tkanivách, sa rozlišujú dva typy regenerácie - fyziologické a reparatívne. Fyziologická regenerácia sa prejavuje vo forme hypertrofie svalových vlákien, ktorá sa prejavuje zvýšením ich hrúbky a dokonca aj dĺžky, zvýšením počtu organel, hlavne myofibríl, ako aj zvýšením počtu jadier, čo sa nakoniec prejavuje zvýšením funkčnej schopnosti svalových vlákien. Použitím rádioizotopovej metódy sa zistilo, že zvýšenie počtu jadier vo svalových vláknach v podmienkach hypertrofie sa dosiahne rozdelením myosatelitových buniek a následným vstupom dcérskych buniek do myosymplastov.
Zvýšenie počtu myofibríl sa dosiahne syntézou aktínových a myozínových proteínov pomocou voľných ribozómov a následným zostavením týchto proteínov do aktínových a myozínových myofilických vlákien paralelne so zodpovedajúcimi vláknami sarkomérov. V dôsledku toho dochádza najprv k zahusťovaniu myofibríl a potom k ich štiepeniu a tvorbe dcérskych myofibríl. Okrem toho nie je možné vytvárať nové aktínové a myozínové myofilické vlákna paralelne, ale s predchádzajúcimi myofibrilmi, čím sa dosiahne ich predĺženie. Sarkoplazmatické retikulum a T-tubuly v hypertrofickom vlákne sa tvoria v dôsledku proliferácie predchádzajúcich prvkov. V niektorých typoch svalový tréning môže sa tvoriť prevažne červený typ svalového vlákna (v štýloch) alebo biely typ svalového vlákna (v sprinteroch). Hypertrofia svalových vlákien spojená s vekom sa intenzívne prejavuje nástupom motorickej aktivity tela (1 až 2 roky), čo je primárne dôsledkom zvýšenej nervovej stimulácie. V senilnom veku, ako aj v podmienkach nízkej svalovej záťaže, dochádza k atrofii špeciálnych a všeobecných organel, riedeniu svalových vlákien a poklesu ich funkčných schopností.
Reparatívna regenerácia sa vyvíja po poškodení svalových vlákien. Metóda regenerácie závisí od veľkosti defektu. Pri významnom poškodení svalového vlákna sú myozatelity v oblasti poškodenia a v priľahlých oblastiach disinhibované, intenzívne proliferujú a potom migrujú do oblasti defektu svalového vlákna, kde sa usporiadajú a tvoria myotubu. Následná diferenciácia myotube vedie k dokončeniu defektu a obnoveniu integrity svalového vlákna. V podmienkach malého defektu vo svalovom vlákne na jeho koncoch, v dôsledku regenerácie intracelulárnych organel, sa tvoria svalové obličky, ktoré rastú smerom k sebe a potom sa zlúčia, čo vedie k uzavretiu defektu. Reparatívnu regeneráciu a obnovenie integrity svalových vlákien je však možné vykonať za určitých podmienok: po prvé, pri zachovanej motorickej inervácii svalových vlákien a po druhé, ak prvky spojivového tkaniva (fibroblasty) nepatria do oblasti poškodenia. Inak sa v mieste defektu svalových vlákien vyvinie jazva spojivového tkaniva.
Sovietsky vedec A.N. Študent preukázal možnosť autotransplantácie tkaniva kostrového svalstva a dokonca aj celých svalov za určitých podmienok:
· Mechanické mletie svalového tkaniva transplantátu s cieľom dezinhibovať satelitné bunky a ich následnú proliferáciu;
· Umiestnenie rozdrveného tkaniva do fasciálneho lôžka;
· Lemovanie vlákniny motorického nervu k rozdrvenému štepu;
· Prítomnosť kontraktívnych pohybov svalových antagonistov a synergistov.
Anatomicky majú novorodenci všetky kostrové svaly, ale z hľadiska telesnej hmotnosti tvoria iba 23% (u dospelých 44%). Množstvo svalovej vlákniny vo svaloch je rovnaké ako u dospelých. Mikroštruktúra svalových vlákien je však odlišná: vlákna majú menší priemer, majú viac jadier. Ako rastie, dochádza k zhrubnutiu a predĺženiu vlákien. Je to kvôli zhrubnutiu myofibríl, ktoré tlačia jadro na perifériu. Veľkosť svalových vlákien sa stabilizuje o 20 rokov.
Svaly u detí sú pružnejšie ako u dospelých. tj skrátiť rýchlejšie pri sťahovaní a predĺžiť pri relaxácii. Vzrušivosť a labilita svalov novorodencov je nižšia ako u dospelých, ale s vekom sa zvyšuje. U novorodencov, dokonca aj vo sne, sú svaly v tónovom stave. K rozvoju rôznych svalových skupín dochádza nerovnomerne. Po 4-5 rokoch sú svaly predlaktia rozvinutejšie, svaly ruky zaostávajú vo vývoji. K zrýchlenému otepľovaniu svalov ruky dochádza za 6-7 rokov. Okrem toho sa extenzory vyvíjajú pomalšie ako flexory. S vekom sa pomer svalového tonusu mení. V ranom detstve zvýšený tón svalov ruky, extenzory bedra atď. Postupne sa normalizuje distribúcia tónov.
Srdce ako orgán sa vyznačuje schopnosťou regenerácie regeneratívnou hypertrofiou, pri ktorej je hmota orgánu obnovená, ale tvar zostáva narušený. Podobný jav sa pozoruje po infarkte myokardu, keď sa hmota srdca môže zotaviť ako celok, zatiaľ čo v mieste poškodenia sa vytvorí jazva spojivového tkaniva, ale hypertrofia orgánov, t.j. formulár je rozbitý. Vyskytuje sa nielen zväčšenie veľkosti kardiomyocytov, ale aj proliferácia hlavne v predsieňach a ušiach srdca.
Už skôr sa verilo, že diferenciácia kardiomyocytov je ireverzibilný proces spojený s úplnou stratou schopnosti týchto buniek sa deliť. Na súčasnej úrovni však početné údaje ukazujú, že diferencované kardiomyocyty sú schopné syntézy DNA a mitózy. Vo výskumných prácach P.P. Rumyantsev a jeho študenti ukázali, že po experimentálnom infarkte myokardu ľavej komory sa 60-70% predsieňových kardiomyocytov vráti do bunkového cyklu, počet polyploidných buniek sa zvyšuje, ale to nekompenzuje poškodenie myokardu.
Zistilo sa, že kardiomyocyty sú schopné mitotického delenia (vrátane buniek vodivého systému). V myokarde srdca je zvlášť veľa mononukleárnych polyploidných buniek s 16-32-násobným obsahom DNA, ale existujú aj binárne kardiomyocyty (13-14%), najmä oktopoid.
V procese regenerácie srdcového svalového tkaniva sa kardiomyocyty podieľajú na procese hyperplázie a hypertrofie, zvyšuje sa ich ploidia, ale úroveň proliferácie buniek spojivového tkaniva v oblasti poškodenia je 20-40 krát vyššia. Vo fibroblastoch je aktivovaná syntéza kolagénu, v dôsledku čoho dochádza k oprave zjazvením defektu. Biologická prezentácia takejto adaptívnej reakcie spojivového tkaniva sa vysvetľuje životnou dôležitosťou srdcového orgánu, pretože oneskorenie pri uzavretí defektu môže viesť k smrti.
Verilo sa, že u novorodencov a pravdepodobne aj v ranom detstve, keď stále pretrvávajú schopnosti delenia kardiomyocytov, sú regeneračné procesy sprevádzané zvýšením počtu kardiomyocytov. Okrem toho sa u dospelých fyziologická regenerácia uskutočňuje v myokarde hlavne intracelulárnou regeneráciou bez zvýšenia počtu buniek, t.j. v myokarde dospelých nedochádza k proliferácii kardiomyocytov. Nedávno sa však získal dôkaz, že v zdravom ľudskom srdci je 14 z milióna myocytov v stave mitózy, čo vedie k cytotómii, t.j. počet buniek nie je významný, ale zvyšuje sa.
Použitie moderných metód bunkovej biológie v klinických a experimentálnych štúdiách nám umožnilo objasniť bunkové a molekulárne mechanizmy poškodenia a regenerácie myokardu. Obzvlášť zaujímavý je dôkaz, že v perinecrotických oblastiach a vo funkčne preťaženom srdci prebieha syntéza embryonálnych proteínov a peptidov myokardu, ako aj proteínov syntetizovaných počas bunkového cyklu. To potvrdzuje podobnosť medzi mechanizmami regenerácie a normálnou ontogenézou.
Ukázalo sa tiež, že diferencované kardiomyocyty v kultúre sú schopné aktívneho mitotického delenia, čo sa dá vysvetliť nie úplnou stratou, ale potlačením schopnosti kardiomyocytov vrátiť sa do bunkového cyklu.
Dôležitou úlohou teoretickej a praktickej kardiológie je vývoj spôsobov, ako stimulovať obnovu poškodeného myokardu, t. indukcia regenerácie myokardu a zníženie jazvy spojivového tkaniva. Jedna z oblastí výskumu poskytuje možnosť prenosu regulačných génov, ktoré premieňajú zjazvené fibroblasty na myoblasty alebo transfekcie na kardiomyocyty génov, ktoré kontrolujú rast nových buniek. Ďalším smerom je prenos do oblasti poškodenia fetálnych kostrových a myokardiálnych buniek, ktoré by sa mohli podieľať na obnove srdcového svalu. Uskutočňujú sa aj experimenty v oblasti transplantácie kostrového svalstva v srdci, ktoré ukazujú tvorbu oblastí sťahujúcich tkanivo v myokarde a zlepšujú funkčné indexy myokardu. Liečba s použitím rastových faktorov, ktoré majú priame aj nepriame účinky na poškodený myokard, napríklad zlepšenie angiogenézy, môže byť sľubná.
Hladké svalové tkanivo
Pôvodom sú tri skupiny hladkých (alebo pruhovaných) svalových tkanív - mezenchymálne, epidermálne a nervové.
Svalové tkanivo mezenchymálneho pôvodu
Histogenézy. Kmeňové bunky a progenitorové bunky hladkého svalstva, ktoré už boli stanovené, migrujú na miesta nosných orgánov. Odlíšia sa tým, že syntetizujú zložky matrice a kolagén základnej membrány, ako aj elastín. V konečných bunkách (myocytoch) je syntetická schopnosť znížená, ale úplne nezmizne.
Štruktúrna a funkčná jednotka hladkého alebo nezúženého svalového tkaniva je bunka hladkého svalstva alebo hladký myocyt je vretenovitá bunka dlhá 20 až 500 μm, šírka 5 až 8 μm. Jadro bunky je v tvare tyčinky, umiestnené v jej centrálnej časti. Keď sa myocyt sťahuje, jeho jadro sa ohýba a dokonca sa krúti. Organely všeobecného významu, medzi ktorými je veľa mitochondrií, sú sústredené v cytoplazme v blízkosti pólov jadra. Golgiho aparát a granulované endoplazmatické retikulum sú slabo vyvinuté, čo naznačuje nízku aktivitu syntetických funkcií. Ribozómy sa väčšinou nachádzajú voľne.
Aktínové vlákna tvoria v cytoplazme trojrozmernú sieť, predĺženú hlavne pozdĺžne alebo skôr šikmo pozdĺžne. Konce nekonečných vlákien sú navzájom spojené a k plazmolemme pomocou špeciálnych zosieťovacích proteínov. Tieto oblasti sú jasne viditeľné v elektrónových mikrografoch ako husté telá.
Myozínové vlákna sú v depolymerizovanom stave. Myozínové monoméry sa nachádzajú vedľa aktínových vlákien. Signál kontrakcie zvyčajne prichádza cez nervové vlákna. Mediátor, ktorý vyčnieva zo svojich terminálov, mení stav plazmolémie. Tvorí vniknutia - jaskyňu, v ktorej sú koncentrované ióny vápnika. Jaskyne sa privádzajú do cytoplazmy vo forme bublín (tu sa z bublín uvoľňuje vápnik). To znamená tak polymerizáciu myozínu, ako aj interakciu myozínu s aktínom. Aktínové vlákna sa posúvajú k sebe, husté škvrny sa spoja, sila sa prenesie na plazmatickú membránu a celá bunka sa skráti. Po zastavení signálov z nervového systému sa z jaskyne evakuujú vápenaté ióny, myozín sa depolymerizuje a myofibrily sa rozpadnú. Komplexy aktín-myozín teda existujú v hladkých myocytoch iba počas obdobia kontrakcie.
Hladké myocyty sa nachádzajú bez viditeľných medzibunkových priestorov a sú oddelené bazálnou membránou. V niektorých oblastiach sa v ňom vytvárajú „okná“, takže sa zbližuje plazma- léma susedných myocytov. Vytvárajú sa tu prípady a medzi bunkami vznikajú nielen mechanické, ale aj metabolické väzby. Na vrchnej časti „krytov“ z bazálnej membrány medzi myocytmi sú elastické a retikulárne vlákna, ktoré kombinujú bunky do jedného tkanivového komplexu. Reticulárne vlákna prenikajú do trhlín na koncoch myocytov, fixujú sa tam a prenášajú silu bunkovej kontrakcie na celé spojenie.
Regenerácie. Fyziologická regenerácia tkaniva hladkého svalstva sa prejavuje v podmienkach zvýšeného funkčného zaťaženia. To je najzreteľnejšie vidieť na svalovej membráne maternice počas tehotenstva. Takáto regenerácia sa neuskutočňuje ani tak na úrovni tkanív ako na bunkovej úrovni: rastú myocyty, v cytoplazme sa aktivujú syntetické procesy, zvyšuje sa počet myofilamentov (hypertrofia pracovných buniek). Avšak bunková proliferácia (t.j. hyperplázia) nie je vylúčená.
Ako súčasť orgánov sa myocyty kombinujú do zväzkov, medzi ktorými sú tenké vrstvy spojivového tkaniva. Do týchto vrstiev sú tkané retikulárne a elastické vlákna obklopujúce myocyty. Cez medzivrstvy prechádzajú krvné cievy a nervové vlákna. Konce posledne uvedených nekončia priamo na myocytoch, ale medzi nimi. Preto sa po príchode nervového impulzu mediátor šíri difúzne a vzrušuje mnoho buniek naraz. Hladké svalové tkanivo mezenchymálneho pôvodu je zastúpené hlavne v stenách krvných ciev a mnohých tubulárnych vnútorných orgánoch a tiež tvorí jednotlivé malé svaly.
Hladké svalové tkanivo v konkrétnych orgánoch má nerovnaké funkčné vlastnosti. Je to spôsobené skutočnosťou, že na povrchu orgánov existujú rôzne receptory pre špecifické biologicky aktívne látky. Preto pri mnohých liekoch nie je ich reakcia rovnaká.
Hladké svalové tkanivo epidermálneho pôvodu
Myoepiteliálne bunky sa vyvíjajú z epidermálneho zárodku. Nachádzajú sa v potných, mliečnych, slinných a slzných žľazách a majú spoločné prekurzory so žľazovými sekrečnými bunkami. Myoepiteliálne bunky sú priamo spojené s vlastným epitelom a majú s nimi spoločnú bazálnu membránu. Počas regenerácie sa tieto a ďalšie bunky obnovia z bežných nízko diferencovaných prekurzorov. Väčšina myoepiteliálnych buniek má tvar hviezdy. Tieto bunky sa často nazývajú kôšovité: ich procesy pokrývajú koncové úseky a malé kanály žľazy. Jadro a organely všeobecného významu sa nachádzajú v tele bunky a kontraktívne aparáty, usporiadané rovnako ako v bunkách svalového tkaniva mezenchymálneho typu, sa nachádzajú v procesoch.
Hladké svalové tkanivo nervového pôvodu
Myocyty tohto tkaniva sa vyvíjajú z buniek nervového základu ako súčasť vnútornej steny optického pohárika. Telá týchto buniek sú umiestnené v epiteli zadného povrchu dúhovky. Každá z nich má proces, ktorý prechádza do hrúbky dúhovky a leží rovnobežne s jej povrchom. Pritom existuje kontraktilný aparát, usporiadaný rovnakým spôsobom ako vo všetkých hladkých myocytoch. V závislosti od smeru procesov (kolmých alebo rovnobežných s okrajom zornice) tvoria myocyty dva svaly - zúženie a zväčšenie zrenice.
záver
Ako už bolo uvedené, svalové tkanivo je skupina telových tkanív rôzneho pôvodu, zjednotená na princípe kontraktility: pruhované (kostrové a srdcové), hladké a špecializované kontraktívne tkanivá - epiteliálne svalové a neurogliálne, ktoré sú súčasťou dúhovky oka.
Pruhované tkanivo kostrového svalstva vzniká z myotómov, ktoré tvoria prvky segmentovaného mezodermu - somitov.
Hladké svalové tkanivo u ľudí a stavovcov sa vyvíja ako súčasť derivátov mezenchýmu, ako aj vnútorného tkaniva. Všetky svalové tkanivá sa však vyznačujú podobnou izoláciou v embryonálnom embrye vo forme vretienkovitých buniek - svalovotvorných buniek alebo myoblastov.
Kontrakcia svalových vlákien zahŕňa skracovanie myofibríl v každom sarkomere. Hrubé (myozínové) a tenké (aktínové) vlákna v uvoľnenom stave, spojené iba koncovými úsekmi, vykonávajú v čase kontrakcie kĺzavé pohyby smerom k sebe. Uvoľňovanie energie potrebné na zníženie energie nastáva v dôsledku premeny ATP na ADP pod vplyvom myozínu. Enzymatická aktivita myozínu sa prejavuje za podmienky optimálneho obsahu Ca2 +, ktorý sa hromadí v sarkoplazmatickom retikule.
Referencie
1. Histológia. Editoval Yu.I. Afanasyeva, N.A. Yurina. M.: „Medicine“, 1999
2. R. Eckert, D. Rendel, J. Augustine „Fyziológia zvierat“ - 1 diel M .: „Mir“, 1981
3. K.P. Ryabov „Histológia so základmi embryológie“ Minsk: „Vyššia škola“, 1990
4. Histológia. Editoval Ulumbekov, prof. YA Chelisheva. M .: 1998
5. Histológia. Editoval V.G. Eliseev. M.: „Medicine“, 1983
Publikované na Allbest.ru
...Podobné dokumenty
Štrukturálne vlastnosti svalového tkaniva. Štúdium mechanizmu svalovej kontrakcie a prenosového excitačného aparátu. Histogenéza a regenerácia svalového tkaniva. Princípy kontraktívnych, vodivých a sekrečných kardiomyocytov srdcového svalu.
cheat sheet pridaný 14/14/2010
Štúdium typov a funkcií rôznych ľudských tkanív. Úlohy histologickej vedy, ktorá študuje štruktúru tkanív živých organizmov. Štrukturálne vlastnosti epitelového, nervového, svalového tkaniva a tkanív vnútorného prostredia (spojivové, kostrové a tekutinové).
prezentácia pridaná 11. 8. 2013
Štúdium štruktúrnych vlastností živočíšnych tkanív, fungovania a odrôd. Uskutočnenie štúdie charakteristického rysu štruktúry spojivového a nervového tkaniva. Štruktúra plochého, kubického, riasnatého a žľazového epitelu. Druhy svalového tkaniva.
prezentácia pridaná 2. 8. 2015
Všeobecné vlastnosti a vekové vlastnosti chrupavkového tkaniva. Druhy chrupavky a kostného tkaniva. Všeobecné vlastnosti a vlastnosti kostného tkaniva súvisiace s vekom. Charakteristiky štruktúry svalového tkaniva v detstve a v starobe. Kostrové svalové tkanivo.
prezentácia, pridané 02/07/2016
Klasifikácia tkanív, typy epitelových tkanív, ich štruktúra a funkcie. Podporná, trofická a ochranná funkcia spojivového tkaniva. Funkcie nervového a svalového tkaniva. Pojem orgány a orgánové systémy, ich individuálne, pohlavie, vekové rozdiely.
abstrakt, pridané 09/11/2009
Epitelové tkanivo, jeho regeneračná schopnosť. Spojivové tkanivá zapojené do udržiavania homeostázy vnútorného prostredia. Bunky útulku a lymfy. Pružné a srdcové svalové tkanivo. Funkcie nervových buniek a tkanív živočíšnych organizmov.
abstrakt, pridané 16. januára 2015
Štúdium druhov živočíšneho tkaniva, ako aj funkcií, ktoré vykonávajú. Štrukturálne vlastnosti skupín tkanív epitelu, spojiva, svalov a nervov. Určenie umiestnenia každej skupiny a dôležitosť pre život zvieraťa.
prezentácia, pridané 18.10.2013
Všeobecná koncepcia a typy vibrácií. Charakterizácia procesov napätia (stláčania), strihu, ohybu, krútenia. Mechanické vlastnosti kostí a cievneho tkaniva. Špecifickosť svalového tkaniva, hlavné spôsoby práce svalov - izometrické a izotonické.
kontrolné práce, pridané 19.03.2014
Bunka ako hlavná stavebná jednotka tela. Opis jeho štruktúry, životných a chemických vlastností. Štruktúra a funkcie epitelového a spojivového, svalového a nervového tkaniva. Orgány a zoznam systémov ľudských orgánov, ich účel a funkcie.
prezentácia pridaná 19. 4. 2012
Fyziológia a biochémia svalovej aktivity ako dôležitej zložky metabolizmu v tele. Druhy svalového tkaniva a podľa toho svaly, ktoré sa líšia štruktúrou svalových vlákien, povahou inervácie. Účinok pohybovej aktivity rôznej intenzity.
Kapitola 9. MUSCULAR TISSUES
Kapitola 9. MUSCULAR TISSUES
Svalové tkanivo (textus muscularis)nazývané látky, ktoré sa líšia štruktúrou a pôvodom, ale majú podobnú schopnosť znižovať sa. Poskytujú pohyb v celom tele tela, jeho častiach a pohybe orgánov vo vnútri tela (srdce, jazyk, črevá atď.).
Bunky mnohých tkanív majú schopnosť sťahovať sa so zmenou tvaru, ale vo svalových tkanivách sa táto schopnosť stáva hlavnou funkciou.
9.1. VŠEOBECNÉ MORPHOFUNKČNÉ CHARAKTERISTIKY A KLASIFIKÁCIA
Hlavnými morfologickými znakmi prvkov svalového tkaniva sú pretiahnutý tvar, prítomnosť pozdĺžne umiestnených myofibríl a myofilamentov - špeciálne organely, ktoré zabezpečujú kontraktilitu, lokalizáciu mitochondrií v blízkosti kontraktilných prvkov, prítomnosť inklúzií glykogénu, lipidov a myoglobínu.
Špeciálne kontraktilné organely - myofily alebo myofibrily, poskytujú kontrakciu, ktorá nastane, keď s nimi interagujú dva hlavné fibrilárne proteíny - aktín a myozín, s povinnou účasťou iónov vápnika. Mitochondrie dodávajú týmto procesom energiu. Zásobu energetických zdrojov tvoria glykogén a lipidy. Myoglobín je proteín, ktorý zaisťuje viazanie kyslíka a vytváranie jeho rezervy v čase kontrakcie svalov, keď sú krvné cievy stlačené (prietok kyslíka prudko klesá).
Klasifikácia.Klasifikácia svalového tkaniva je založená na dvoch zásadách - morfofunkčnom a histogenetickom. V súlade s morfologickým a funkčným princípom sa svalové tkanivo v závislosti od štruktúry organel kontrakcie delí na dve podskupiny.
Prvá podskupina- priečne pruhované (priečne pruhované) svalové tkanivo (textus muscularis striatus).V cytoplazme ich prvkov, myozínové vlákna
ste neustále polymerizovaní, vytvárajte neustále existujúce myofibrily s aktínovými vláknami. Tie sú organizované v charakteristických komplexoch - sarcomeres.V susedných myofibrilách sú štruktúrne podjednotky sarkomérov umiestnené na rovnakej úrovni a vytvárajú sa priečne pruhovanie.
Druhá podskupina- hladké (neviazané) svalové tkanivo (textus muscularis nonstriatus).Tieto tkanivá sa vyznačujú skutočnosťou, že bez kontrakcie sa myozínové vlákna depolymerizujú. V prítomnosti vápenatých iónov polymerizujú a interagujú s aktínovými vláknami. Myofibrily vytvorené v tomto prípade nemajú priečne pruhovanie: so špeciálnymi farbami sú reprezentované (hladkými) vláknami rovnomerne zafarbené po celej dĺžke.
V súlade s histogénnym princípom sa svalové tkanivá a svalové prvky v závislosti od zdrojov vývoja (embryonálna primordia) delia na: somatické (myotóm), coelomické (z myoepikardiálnej doštičky viscerálneho splanchnotómového listu), mezenchymálne (z desmálneho zárodku v mezenchýme (nervové)), nervové nervová trubica), epidermálna (z kožnej ektodermy az predchordálnej doštičky).
9.2. STRIPPED MUSCULAR TISSUES
Existujú dve hlavné odrody priečne pruhovaných (priečne pruhovaných) tkanív - kostrové (myotóm) a srdcové (coelomické).
9.2.1. Kostrové svalové tkanivo
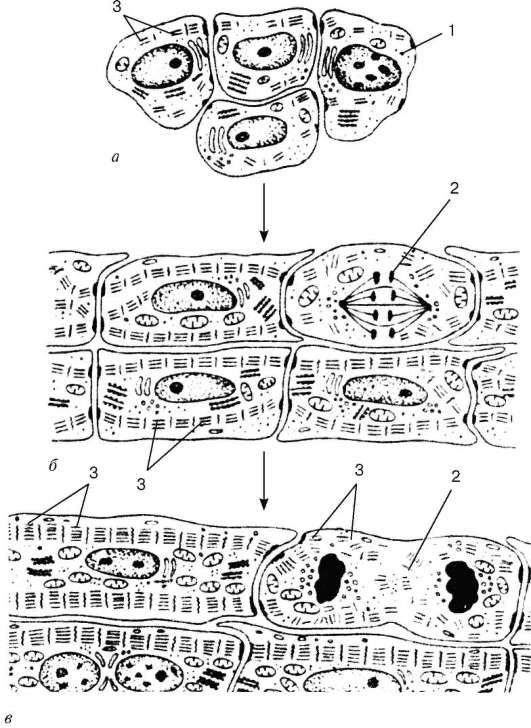
Histogenézy.Zdroj vývoja prvkov kostrového (somatického) pruhovaného svalového tkaniva (textus muscularis striatus sceletalis)sú kmeňové bunky myotómov - promioblasty. Niektoré z nich sa na mieste diferencujú a podieľajú sa na tvorbe tzv. Autochtónnych svalov. Ostatné bunky migrujú z myotómov do mezenchýmu. Sú už určené, aj keď navonok sa nelíšia od iných mezenchýmových buniek. Ich diferenciácia pokračuje v miestach, kde sú položené ďalšie svaly tela. Počas diferenciácie vznikajú dve bunkové línie. Bunky jednej z línií sa spájajú a vytvárajú pretiahnuté symptómy - svalové trubice (myotubes). V nich je diferenciácia špeciálnych organel - myofibríl (obr. 9.1). V súčasnosti je v myotubách zaznamenaný dobre vyvinutý granulárny endoplazmatický retikulum. Myofibrily sa najskôr umiestnia pod plazmolémom a potom vyplnia väčšinu myotrubíc. Naopak, jadrá z centrálnych oddelení sú presunuté na perifériu. Bunkové centrá a mikrotubuly úplne vymiznú. Granulárne endo
Obr. 9.1.Histogenéza tkaniva kostrového svalstva (podľa A. A. Klishov):
a- promioblasty; b- myosymplast; v- svalová trubica; g- zrelý sval
vlákno. 1 - myosatelit; 2 - jadro myosymplastov; 3 - myofibrily
plazmová sieť je významne znížená. Takéto definitívne štruktúry sa nazývajú myosymplasty.
Bunky druhej línie zostávajú nezávislé a diferencujú sa na myosatelitné bunky. Tieto bunky sa nachádzajú na povrchu myosymplastov. Myosatelitné bunky, množiace sa, sa spájajú s myosymplastami, a tak sa podieľajú na vytvorení optimálneho jadra
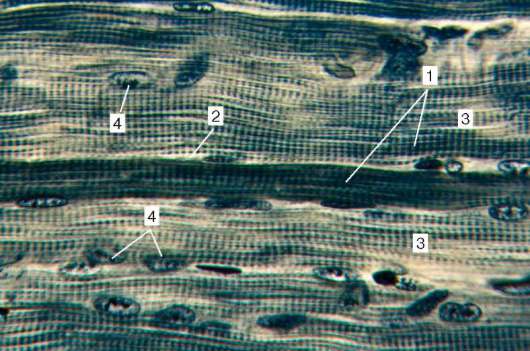
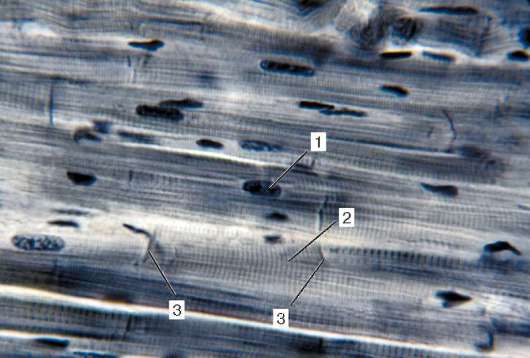
Obr. 9.2.Štruktúra pruhovaného svalového tkaniva (mikrograf):
1 - svalové vlákna; 2 - sarkolemma; 3 - sarkoplazma a myofibrily; 4 - jadrá
myosymplast. Farbenie - hematoxylín železa
sarkoplazmatický vzťah potrebný na syntézu špecifických proteínov symplastov.
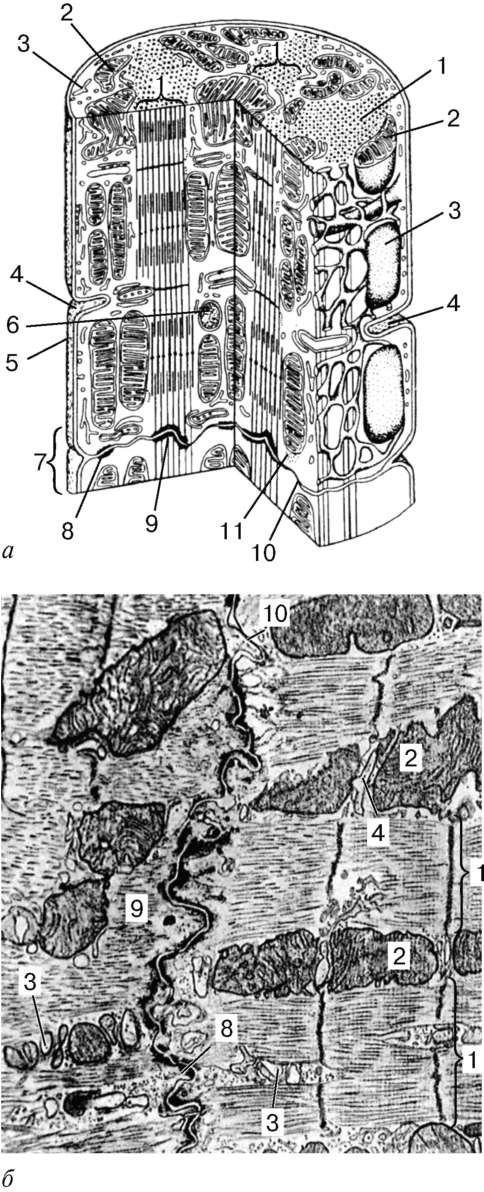
Štruktúra.Hlavnou štrukturálnou jednotkou kostrového svalového tkaniva je svalové vlákno pozostávajúce z myosymplastov a myosatelocytov pokryté spoločnou bazálnou membránou (obr. 9.2-9.4). Dĺžka celého vlákna sa môže merať v centimetroch s hrúbkou 50 až 100 mikrónov. Komplex pozostávajúci z plazmolémie myosymplastov a bazálnej membrány sa nazýva sarkolemma.
Štruktúra myosymplastov.Myosymplast má mnoho podlhovastých jadier, ktoré sa nachádzajú priamo pod plazmolémom. Ich počet v jednom simplaste môže dosiahnuť niekoľko desiatok tisíc (pozri obr. 9.2). Organe všeobecného významu - Golgiho komplex a malé fragmenty agranulárneho endoplazmatického retikula - sa nachádzajú na póloch jadier. Myofibrily vyplňujú hlavnú časť myosymplastov a sú umiestnené pozdĺžne (pozri obrázok 9.3).
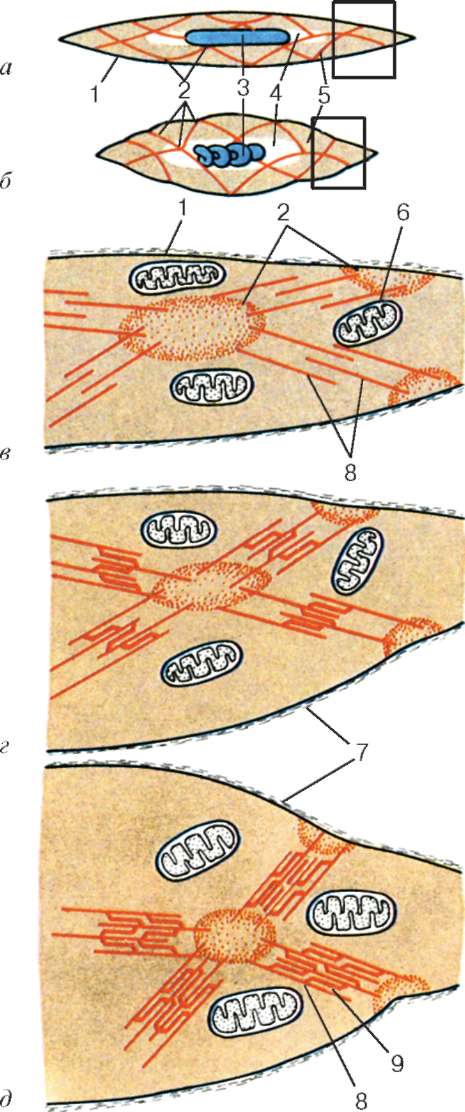
inokomma- štrukturálna jednotka myofibríl. Každý myofibril má priečne tmavé a ľahké disky s nerovnakým lomom (anizotropné A-disky a izotropné I-disky). Každý myofibril je obklopený slučkami agranulárneho endoplazmatického retikula - sarkoplazmatického retikula - pozdĺžne umiestneného a medzi anastomózou. Susedné sarkoméry majú spoločnú štruktúru hraníc - čiaru Z (obr. 9.5). Je postavený vo forme siete proteínových fibrilárnych molekúl, medzi ktorými hrá dôležitú úlohu alfa-aktinín. Konce aktínových vlákien sú pripojené k tejto sieti. Zo susedných Z-línií sú aktínové vlákna nasmerované do stredu sarkoméry, ale nedosahujú jej stred. Aktínové vlákna kombinované s líniou Z a myozínovými vláknami
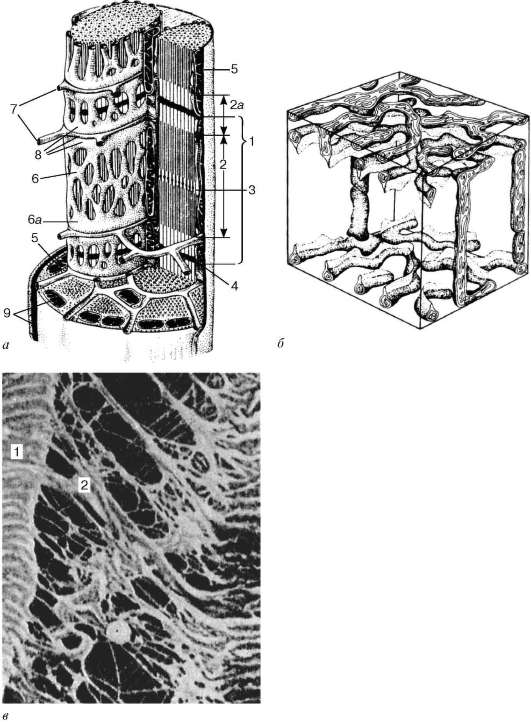
Obr. 9.3.Schéma ultramikroskopickej štruktúry myosymplastov (podľa R. V. Krstika, so zmenami) (a): 1 - sarkomér; 2 - anizotropný disk (dráha A); 2a - izotropný disk (dráha I); 3 - čiara M (mezofragma) uprostred anizotropného disku; 4 - čiara Z (telophragm) uprostred izotropného disku; 5 - mitochondrie; 6 - sar-koplazmatické retikulum; 6a - posledná nádrž; 7 - priečna rúrka (T-trubica); 8 - triáda; 9 - sarkolemma; b- Priestorové usporiadanie mitochondrií v symplaste. Horná a dolná rovina obrázku sú obmedzené anizotropným diskom sarko-mierky (podľa L. E. Bakeeva, V. P. Skulachev, Yu. S. Chentsov); v- endomýzium. Rastrovací elektrónový mikrograf, zväčšenie 2600 (príprava Yu. A. Khoroshkov): 1 - svalové vlákna; 2 - kolagénové vlákna
fibrilárne neroztiahnuteľné molekuly nebulínu. Uprostred tmavého disku sarkoméry je sieť vyrobená z myomyozínu. V priereze tvorí čiaru M. V uzloch tejto M-línie sú konce myozínových filamentov fixované. Ich ďalšie konce sú nasmerované k Z-čiaram a
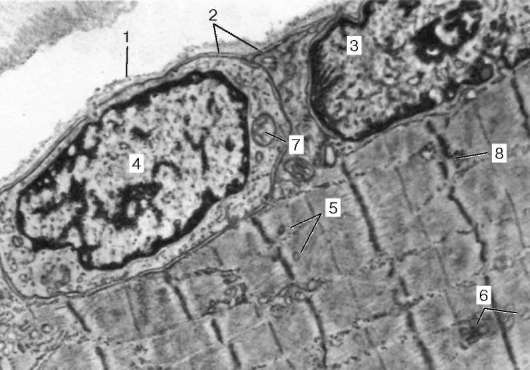
Obr. 9.4.Povrch myosymplastov a myosatelitov. Elektrónový mikrograf, zväčšenie 10 000 (príprava V. L. Goryachkiny a S. L. Kuznetsovej): 1 - suterénna membrána; 2 - plazmoléma; 3 - jadro myosymplasty; 4 - jadro myos Tellitocytu; 5 - myofibrily; 6 - tubuly agranulárnej endoplazmatickej (sarkoplazmovej) siete; 7 - mitochondrie; 8 - glykogén
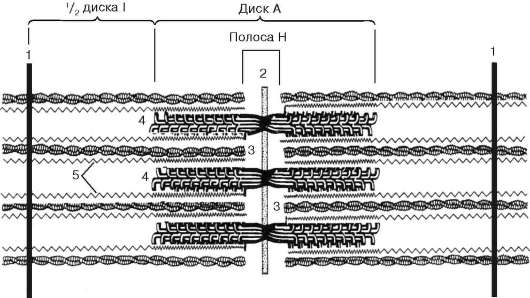
Obr. 9.5.Sarcomere (diagram):
1 - riadok Z; 2 - riadok M; 3 - aktínové vlákna; 4 - myozínové vlákna; 5 - molekuly fibrilárneho titínu (podľa B. Alberts, D. Bray, J. Lewis a kol., V znení zmien a doplnení)
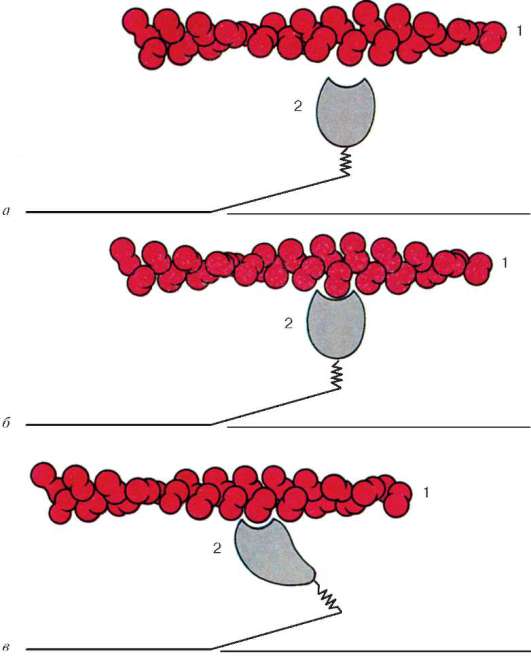
Obr. 9.6.Konformačné zmeny, ktoré si vyžadujú vzájomné vytesnenie aktínových a myozínových vlákien:
ac- postupné zmeny v priestorových vzťahoch. 1 - aktín; 2 - hlava molekuly myozínu (podľa B. Alberts, D. Bray, J. Lewis a kol., V znení zmien a doplnení)
nachádzajú sa medzi aktínovými vláknami, ale tiež nedosahujú samotné Z-línie. Súčasne sú tieto konce fixované vzhľadom na Z línie pomocou ťahaných gigantických proteínových molekúl titínu.
Molekuly myozínu majú na jednom konci dlhý chvost a dve hlavy. So zvýšením koncentrácie iónov vápnika v oblasti pripojenia hláv (pántová časť) molekula zmení svoju konfiguráciu (Obr. 9.6). Súčasne (od aktínu lokalizovaného medzi myozínovými vláknami) sa myozínové hlavy viažu na aktín (s účasťou pomocných proteínov - tropomyozín a troponín). Potom sa myozínová hlava nakloní a vytiahne spolu s ňou molekulu aktínu smerom k M-línii. Z-línie sa zbiehajú, sarkomér je skrátený.
Alfa-aktinínové siete Z-línií susedných myofibríl sú vzájomne prepojené strednými vláknami. Prichádzajú k vnútornému povrchu plazmatémie a sú fixované vo svojej kortikálnej vrstve, takže sarkoméry všetkých myofibríl sú umiestnené na rovnakej úrovni. Toto, keď sa pozoruje pod mikroskopom, vytvára dojem priečneho pruhu celého vlákna.
Zdrojom vápenatých iónov sú nádrže agranulárneho endoplazmatického retikula. Sú podlhovasté pozdĺž myofibríl blízko každého sarkoméru a tvoria sarkoplazmatické retikulum. Práve v ňom sa vápnikové ióny akumulujú, keď je myosymplast v uvoľnenom stave. Na úrovni Z-línií (v obojživelníkoch) alebo na hranici diskov A a I (u cicavcov) sa tubuly smeru siete menia a sú priečne, tvoriace predĺžené terminálne alebo laterálne (L) nádrže.
Z povrchu do hĺbky myosymplastov tvorí plazmolém dlhé tubuly, ktoré sa rozkladajú priečne (T-tubuly) na úrovni hraníc medzi tmavými a svetlými diskami. Keď myosymplast dostane signál o začiatku kontrakcie, pohybuje sa pozdĺž plazmolému vo forme akčného potenciálu a šíri sa na membránu T-trubice. Pretože táto membrána je blízko membrán sarkoplazmatického retikula, mení sa jej stav, vápnik sa uvoľňuje z nádrží siete a interaguje s aktín-myozínovými komplexmi (sťahujú sa). Keď zmizne akčný potenciál, vápnik sa znova hromadí v tubuloch siete a kontrakcia myofibríl sa zastaví. Energia je potrebná na rozvoj úsilia o zníženie emisií. Uvoľňuje sa prevodom ATP na ADP. Úlohu ATPázy vykonáva myozín. Zdrojom ATP sú hlavne mitochondrie, a preto sa nachádzajú priamo medzi myofibrílmi.
Dôležitú úlohu v aktivite myozymplastov zohrávajú inklúzie myoglobínu a glykogénu. Glykogén slúži ako zdroj energie, potrebný nielen na výkon svalovej práce, ale aj na udržanie tepelnej rovnováhy celého organizmu. Myoglobín viaže kyslík, keď je sval uvoľnený a krv voľne prúdi cez malé krvné cievy. Počas sťahovania svalov sú krvné cievy stlačené a ukladaný kyslík sa uvoľňuje a podieľa sa na biochemických reakciách.
Myosatelitné bunky.Tieto zle diferencované bunky sú zdrojom regenerácie svalového tkaniva. Priliehajú k povrchu myosymplastov, takže ich plazmolémy sú v kontakte (pozri obr. 9.1, 9.4). Myosatelitné bunky sú mononukleárne, ich tmavé jadrá sú oválne a menšie ako v symplastoch. Majú všetky organely všeobecného významu (vrátane bunkového centra).
Druhy svalových vlákien.Rôzne svaly (napríklad orgány) fungujú v rôznych biomechanických podmienkach. Preto svalové vlákna v zložení rôznych svalov majú rôznu silu, rýchlosť a trvanie kontrakcie, ako aj únavu. Aktivita enzýmov v nich je rôzna a sú prítomné v rôznych izomérnych formách. Obsah respiračných enzýmov - glykolytických a oxidačných - sa v nich tiež líši.
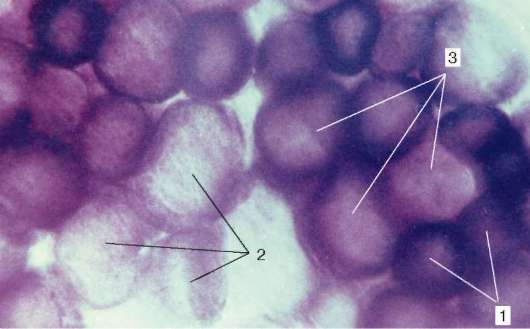
Obr. 9.7.Aktivita sukcinátdehydrogenázy vo svalových vláknach rôznych typov (liek V. F. Chetvergova, spracovanie podľa Nakhlas et al.): 1 - vysoká; 2 - nízka; 3 - stredný
Pomerom myofibríl, mitochondrií a myoglobínu sa rozlišujú biele, červené a stredné vlákna. Podľa funkčných vlastností sa svalové vlákna delia na rýchle, pomalé a stredné, čo je určené molekulovou organizáciou myozínu. Medzi jej izoformami sú dve hlavné - „rýchla“ a „pomalá“. Pri zavádzaní histochemických reakcií sú identifikované aktivitou ATPázy. S týmito vlastnosťami koreluje aj aktivita respiračných enzýmov. V rýchlych vláknach prevažujú glykolytické procesy, sú bohaté na glykogén, majú menej myoglobínu, preto sa nazývajú biele. Naopak, v pomalých vláknach je aktivita oxidujúcich enzýmov vyššia, sú bohatšie na myoglobín a vyzerajú červenšie.
Spolu s bielou a červenou, existujú medziprodukty. Vo väčšine kostrových svalov sú vlákna rôznych histochemických typov mozaikou (obr. 9.7).
Vlastnosti svalových vlákien sa menia s meniacim sa zaťažením - športové, profesionálne, ako aj v extrémnych podmienkach (beztiažové). Keď sa vrátite k normálnym činnostiam, tieto zmeny sú reverzibilné. Pri niektorých chorobách (svalová atrofia, dystrofia, následky denervácie) sa svalové vlákna s rôznymi počiatočnými vlastnosťami menia rôzne. To vám umožní objasniť diagnózu, pri ktorej sa vyšetrujú vzorky biopsie kostrových svalov.
Regenerácie.Jadrá myosymplastov sa nemôžu deliť, pretože v sarkoplazme nie sú žiadne bunkové centrá. Slúžia kambiálne prvky myosatelitné bunky.Keď telo rastie, delia sa a dcérske bunky sa spájajú s myosymplastami. Na konci rastu reprodukcia myosatelitových buniek ustupuje. Po poškodení svalového vlákna z miesta poranenia je zničený a jeho fragment
ste fagocytovaní makrofágmi. Obnovenie tkaniva sa uskutočňuje dvoma mechanizmami: kompenzačnou hypertrofiou samotného symplastu a proliferáciou myosatelitových buniek. V symplastike sa aktivuje granulárne endoplazmatické retikulum a Golgiho komplex. Dochádza k syntéze látok potrebných na obnovenie sarkoplazmy a myofibríl, ako aj k zostaveniu membrán, aby sa obnovila integrita plazmovej lemmy. V tomto prípade sa poškodený koniec myosymplastu zahusťuje a vytvára svalovú obličku. Myosatelitné bunky, ktoré prežívajú v blízkosti lézie, sú rozdelené. Niektoré z nich migrujú do svalovej obličky a integrujú sa do nej, zatiaľ čo iné sa spájajú (rovnako ako myoblasty počas histogenézy) a vytvárajú nové myotubuly, ktoré sa vyvíjajú do svalových vlákien.
9.2.2. Kostrový sval ako orgán
Prenos kontrakčného úsilia na kostru sa uskutočňuje prostredníctvom šliach alebo pripevnením svalov priamo na perioste. Na konci každého svalového vlákna vytvára plazmaéma hlboké úzke hrče. Tenké kolagénové vlákna ich prenikajú zo šľachy alebo periostu. Posledne uvedené sú špirálovo pletené retikálnymi vláknami. Konce vlákien sú nasmerované na bazálnu membránu, vstúpia do nej, otočia sa späť a po výstupe opäť zoradia kolagénové vlákna spojivového tkaniva.
Medzi svalovými vláknami sú tenké vrstvy sypkého vláknitého spojivového tkaniva - endomysium.Kolagénové vlákna vonkajšieho listu bazálnej membrány sú do nej tkané (pozri obr. 9.3, c), čo pomáha spojiť úsilie pri súčasnom znížení počtu myosymplastov. Hustšie vrstvy voľného spojivového tkaniva obklopujú niekoľko svalových vlákien a tvoria sa perimysiuma rozdelenie svalu na zväzky. Niekoľko zväzkov sa spojí do väčších skupín, oddelených hrubšími vrstvami spojivového tkaniva. Nazýva sa spojivové tkanivo obklopujúce povrch svalov epimiziem.
Vaskularizácie.Cievky vstupujú do svalu a šíria sa cez vrstvy spojivového tkaniva a postupne sa stenčujú. Pobočky piateho šiesteho radu tvoria v perimíze arterioly. Kapiláry sa nachádzajú v endomýzii. Prechádzajú svalovými vláknami a navzájom anastomujú. Venuly, žily a lymfatické cievy prechádzajú blízko zásobovacích ciev. Ako je obvyklé, mnoho žírnych buniek v blízkosti ciev sa podieľa na regulácii priepustnosti cievnych stien.
Inervácie.Sval odhalil myelínové efferenty (motorické), aferentné (citlivé) a nemyelínované vegetatívne nervové vlákna. proces nervová bunkaprináša impulz z motorického neurónu miechapobočky v perimisii. Každá z jej vetiev preniká cez bazálnu membránu a tvorí terminály na povrchu simplastu na plazmoléme, podieľajúc sa na organizácii tzv. Motorického plaku (pozri kapitolu 10, obr. 10.18). Po prijatí
Obr. 9.8.Fragment svalového vretena obsahujúci svalové vlákna s jadrovým reťazcom (a) a s jadrovým vakom (b) (schéma podľa G. S. Katinas): 1 - jadrá; 2 - myofibrily (organely všeobecného významu nie sú zobrazené)
acetylcholín je uvoľňovaný z terminálov nervového impulzu - mediátora, ktorý spôsobuje excitáciu (akčný potenciál), ktorý sa odtiaľ šíri prostredníctvom plazmidu myosymplastov.
Každé svalové vlákno je teda inervované nezávisle a je obklopené sieťou hemokapilár, ktoré tvoria komplex nazývaný cASE.
Nazýva sa skupina svalových vlákien inervovaná jedným motorickým neurónom neuromuskulárna jednotka.Svalové vlákna patriace jednej neuromuskulárnej jednotke neležia neďaleko, ale nachádzajú sa medzi mozaikami medzi vláknami patriacimi iným jednotkám.
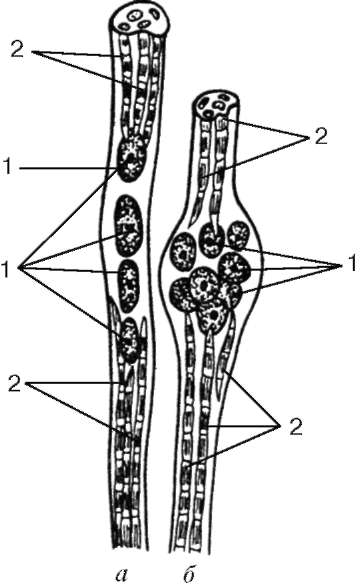
Citlivé nervové zakončenia sa nenachádzajú na pracovných (extrafusal) svalových vláknach, ale sú spojené so špecializovanými svalovými vláknami v tzv. Svalových svaloch.
tenakh (s intrafuzálnymi svalovými vláknami), ktoré sa nachádzajú v perimisiu.
Infúzne svalové vlákna.Vnútrofúzne svalové vlákna vretien sú oveľa tenšie ako pracovníci. Existujú dva typy: vlákno s jadrovým vreckom a vlákno s jadrovým reťazcom (obr. 9.8). Jadrá v týchto a ostatných sú zaoblené a umiestnené v hrúbke symplastiky a nie na jej povrchu. Vo vláknach s jadrovým vreckom tvorí jadro symplastiky zhluky v zosilnenej strednej časti. Vo vláknach s jadrovým reťazcom v strednej časti symplastu nevzniká zahusťovanie, jadrá tu ležia jeden po druhom. Organely všeobecného významu sa nachádzajú v blízkosti zhlukov jadier.
Myofibrily sú umiestnené na koncoch symplastov. Vlákno Sarcolemma sa pripája k kapsule neuromuskulárneho vretena, ktoré pozostáva z hustého vláknitého spojivového tkaniva. Každé svalové vlákno vretena je špirálovito prepletené terminálom senzorického nervového vlákna. V dôsledku redukcie alebo relaxácie pracovných svalových vlákien sa mení napätie kapsuly spojivového tkaniva vretena, respektíve sa mení tón intrafúznych svalových vlákien. V dôsledku toho sú okolo nich vzrušené citlivé nervové zakončenia a v terminálnej oblasti sa objavujú aferentné nervové impulzy. Každý myosymplast má tiež svoj vlastný motorický štítok. Preto sú intrafúzne svalové vlákna neustále v napätí a prispôsobujú sa dĺžke brušného svalu ako celku.
9.2.3. Tkanivo srdcového svalu
Histogenéza a typy buniek.Zdroje rozvoja svalového tkaniva pruhovaného srdcom (textus muscularis striatus cardiacus)- symetrické rezy splanchnotómu viscerálneho listu v krčnej časti embrya - myoepikardiálne platne.Z nich sa diferencujú aj bunky mezoteliomu epikardu. Pôvodné bunky tkaniva srdcového svalu - kardiomyo výbuchy- charakterizované množstvom príznakov: bunky sú sploštené, obsahujú veľké jadro, ľahkú cytoplazmu, chudobné na ribozómy a mitochondrie. V budúcnosti sa vyvíja komplex Golgi, granulovaná endo-plazmatická sieť. Fibrilárne štruktúry sa nachádzajú v kardiomyoblastoch, ale neexistujú žiadne myofibrily. Bunky majú vysoký proliferačný potenciál.
Po sérii mitotických cyklov sa kardiomyoblasty diferencujú na kardiomyocytov,v ktorej začína sarkomogenéza (obr. 9.9). V cytoplazme kardiomyocytov sa zvyšuje počet polysómov a tubulov zrnitého endoplazmatického retikula, hromadí sa glykogénové granule a zvyšuje sa objem aktomyozínového komplexu. Kardiomyocyty sa sťahujú, ale nestrácajú schopnosť ďalej sa množiť a diferencovať. Vývoj kontraktilného aparátu v neskorých embryonálnych a postnatálnych obdobiach nastáva pridaním nových sarkomérov a stratifikáciou novo syntetizovaných myofilamentov.
Diferenciácia kardiomyocytov je sprevádzaná zvýšením počtu mitochondrií, ich distribúciou v póloch jadier a medzi myofibrílami a prebieha paralelne so špecializáciou kontaktných povrchov buniek. Kardiomyocyty tvoria end-to-end, end-to-side kontakty, aby vytvorili srdcové svalové vlákna, a všeobecne, tkanivo je sieťovitá štruktúra. Niektoré kardiomyocyty v raných štádiách kardiomyogenézy sú kontraktívne sekrečné. Následne v dôsledku odlišnej diferenciácie vznikajú „tmavé“ (kontraktilné) a „ľahké“ (vodivé) myocyty, v ktorých zmiznú sekrečné granule, zatiaľ čo zostávajú v predsieňových myocytoch. Vytvára sa teda endokrinný diferencia kardiomyocytov. Tieto bunky obsahujú centrálne lokalizované jadro s dispergovaným chromatínom, jeden alebo dva jadrá. V cytoplazme je dobre vyvinuté granulované endoplazmatické retikulum, diktiómy Golgiho komplexu, v úzkom spojení s prvkami, ktoré sú početné sekrečné granule s priemerom asi 2 um, obsahujúce elektrón-hustý materiál. Následne sa pod sarkolemou nachádzajú sekrečné granule a exocytózou sa vylučujú do medzibunkového priestoru.
Počas histogenézy všeobecne vzniká päť typov kardiomyocytov - pracovné (kontraktilné), sínusové (kardiostimulátor), prechodné, vodivé a tiež sekrečné. Pracovné (kontraktilné) kardiomyocytytvoria ich reťazce (obr. 9.10). Sú skrátené a spôsobujú kontrakciu celého srdcového svalu. Pracovné kardiomyocyty sú schopné
Obr. 9.9.Histogenéza srdcového svalového tkaniva (schéma podľa P.P. Rumyantsev): a- kardiomyocyty v stene srdcovej trubice; b -kardiomyocyty v neskorej embryogenéze; v- kardiomyocyty v postanatálnom období. 1 - kardiomyocyt; 2 - mitoticky sa deliace kardiomyocyty; 3 - myofily a myofibrily
prenášajú navzájom riadiace signály. Sínusové (kardiostimulátory) kardiomyocytyschopný automaticky v určitom rytme zmeniť stav kontrakcie na stav relaxácie. Bunky vnímajú kontrolné signály z nervových vlákien, v reakcii na ktorú menia rytmus kontraktilnej aktivity. Sínusové (kardiostimulátory) kardiomyocyty prenášajú kontrolné signály prechodné kardiomyocyty,a posledne menované na vodivé a pracovné kardiomyocyty. Vedenie kardiomyocytovtvoria reťazce buniek spojených svojimi koncami a sú umiestnené pod endo
Obr. 9.10.Štruktúra srdcového svalového tkaniva (mikrograf). Farbenie - hematoxylín železa:
1 - jadro kardiomyocytov; 2 - reťazec kardiomyocytov; 3 - vložte kolesá
cardo. Prvá bunka v reťazci prijíma kontrolné signály zo sínusových kardiomyocytov a prenáša ich na ďalšie vodivé kardiomyocyty. Bunky, ktoré uzatvárajú reťazec, prenášajú signál cez prechodné kardiomyocyty na pracovníkov. Sekrečné kardiomyocytyvykonávať špeciálnu funkciu. Produkujú peptidový hormón kardiodilatín, ktorý cirkuluje v krvi ako kardionatrín, spôsobuje zníženie hladkých arteriol myocytov, zvýšenie prietoku krvi obličkami a urýchľuje glomerulárnu filtráciu a vylučovanie sodíka. Všetky kardiomyocyty sú pokryté bazálnou membránou.
Štruktúra kontraktívnych (pracovných) kardiomyocytov.Bunky majú podlhovastý (100 - 150 μm) tvar, takmer valcovitý. Ich konce sú navzájom spojené, takže bunkové reťazce tvoria takzvané funkčné vlákna (až 20 mikrónov hrubé). V oblasti kontaktov buniek sa vytvárajú tzv. Vkladacie disky (obr. 9.10). Kardiomyocyty môžu vetviť a vytvárať priestorovú sieť. Ich povrchy sú pokryté bazálnou membránou, do ktorej sú zvonka tkané retikulárne a kolagénové vlákna. Jadro kardiomyocytov (niekedy aj dvoch) je oválne a leží v centrálnej časti bunky. Na póloch jadra sa sústreďuje niekoľko organel všeobecného významu, s výnimkou agranulárneho endoplazmatického retikula a mitochondrie.
Špeciálne organely, ktoré poskytujú kontrakciu, sa nazývajú myofibrily. Sú slabo izolované od seba, môžu sa rozdeliť. Ich štruktúra je podobná štruktúre myofibríl myosymplastov vlákniny kostrového svalstva. Každá mitochondria je umiestnená v celom sarkomere. T-trubice umiestnené na úrovni línie Z sú nasmerované z povrchu plazmovej membrány do hĺbky kardiomyocytov. Ich membrány sú spojené
v kontakte s membránami hladkej endoplazmatickej (sarkoplazmovej) siete. Slučky z nich sú pretiahnuté pozdĺž povrchu myofibríl a majú bočné predĺženia (systémy L), ktoré spolu s T-trubicami tvoria trojicu alebo dyad (obr. 9.11, a). V cytoplazme sú inklúzie glykogénu a lipidov, najmä veľa inklúzií myoglobínu. Mechanizmus kontrakcie kardiomyocytov je rovnaký ako v myosymplastoch.
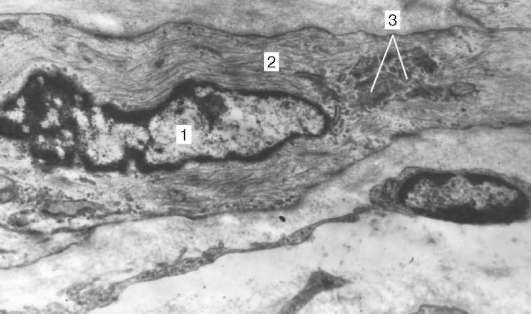
Organizácia kardiomyocytov do tkanív.Kardiomyocyty sú navzájom spojené typom „koniec-koniec“. Tu sa tvoria vkladacie disky: tieto oblasti vyzerajú ako tenké platne s priemerným zväčšením svetelného mikroskopu. Konce kardiomyocytov majú v skutočnosti nerovný povrch, takže výstupky jednej bunky vstupujú do žľabov druhej. Priečne rezy výčnelkov susedných buniek sú vzájomne prepojené interdigitáciami a desmozómami (obr. 9.11, b).
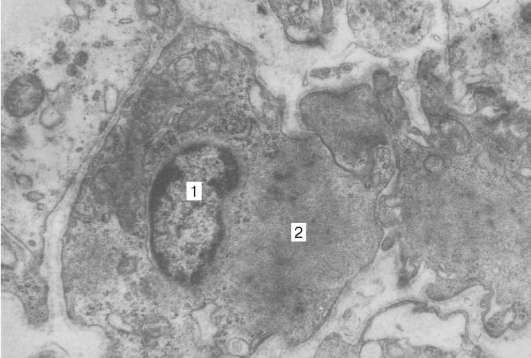
Obr. 9.11.Štruktúra kardiomyocytov: a- schéma (podľa Yu. I. Afanasyev a V. L. Goryachkina); b- elektronický mikrograf vkladacieho disku. Zvýšenie o 20 000. 1 - myofibrily; 2 - mitochondrie; 3 - sarkoskulárna sieť; 4 - T-trubice; 5 - suterénna membrána; 6 - lyzozóm; 7 - vložte disk; 8 - desmozóm; 9 - zóna pripojenia myofibríl; 10 - drážkové kontakty; 11 - glykogén
Ku každému desmozómu zo strany cytoplazmy sa blíži myofibril la, ktorý je na konci fixovaný v komplexe desmo-plak. Teda pri znížení túžby jedného kardiomyocytu sa prenáša na iný. Bočné povrchy výčnelkov kardiomyocytov sú spojené nexusmi (medzerové spoje). To vytvára medzi nimi metabolické väzby a zaisťuje synchronizáciu kontrakcií.
Regenerácie.V histogenéze tkaniva srdcového svalu sa kambium nevyskytuje. Preto regenerácia tkaniva prebieha na základe intracelulárnych hyperplastických procesov. Súčasne je proces polyploidie charakteristický pre kardiomyocyty cicavcov, primátov a ľudí.
vanie. Napríklad u opíc sa až z 50% terminálne diferencovaných kardiomyocytov v jadre stanú tetra- a octoploidmi. Polyploidné kardiomyocyty sa vyskytujú v dôsledku acitokinetickej mitózy, ktorá vedie k multinukleácii. V podmienkach patológie ľudského kardiovaskulárneho systému (reumatizmus, vrodené srdcové defekty, infarkt myokardu, atď.), Intracelulárna regenerácia, polyploidizácia jadier a výskyt viacjadrových kardiomyocytov hrajú dôležitú úlohu pri kompenzácii poškodenia kardiomyocytov.
9.3. HLADKÉ MUSCULÁRNE Tkaniny
Existujú tri skupiny hladkého (neviazaného) svalového tkaniva (textus muscularis nonstriatus)a bunky: mezenchymálne, nervové a myoepiteliálne bunky.
9.3.1. Svalové tkanivo mezenchymálneho pôvodu
Histogenézy.Toto tkanivo je rozdelené na dva typy: viscerálne a vaskulárne. V embryonálnej histogenéze je ťažké rozlíšiť dokonca aj elektrónovú mikroskopiu medzi progenitormi mezenchymálnych fibroblastov a hladkými myocytmi. V zle diferencovaných hladkých myocytoch sa vyvinie granulárne endoplazmatické retikulum, Golgiho komplex. Tenké vlákna sú orientované pozdĺž pozdĺžnej osi bunky. Ako sa vyvíjajú, veľkosť buniek a počet vlákien v cytoplazme sa zvyšuje. Postupne sa zvyšuje objem cytoplazmy obsadenej kontraktilnými vláknami, ich umiestnenie v cytoplazme sa stáva stále viac usporiadaným. Proliferatívna aktivita hladkých myocytov v myogenéze sa postupne znižuje. K tomu dochádza v dôsledku predĺženia trvania bunkového cyklu, výstupu buniek z reprodukčného cyklu a prechodu do diferencovaného stavu. Odlíšia sa tým, že syntetizujú zložky medzibunkovej matrice, kolagén základnej membrány, ako aj elastín. V konečných bunkách (myocytoch) je syntetická schopnosť znížená, ale úplne nezmizne.
Štruktúra a fungovanie buniek.Hladký myocyt je vretenovitá bunka dlhá 20 až 500 mikrometrov, široká 5 až 8 mikrometrov. Jadro je v tvare tyče, umiestnené v jeho stredovej časti. Keď sa myocyty sťahujú, jeho jadro sa ohýba a dokonca sa krúti (Obr. 9.12–9.14).
Štruktúra definitívnych hladkých myocytov (leiomyocytov), \u200b\u200bktoré tvoria vnútorné orgány a steny krvných ciev, má veľa spoločného, \u200b\u200bale zároveň sa vyznačuje heteromorfizmom. Takže v stenách žíl a artérií sa nachádzajú vaječné, v tvare vretena spracované myocyty s dĺžkou 10 až 40 mikrónov, niekedy dosahujúce až 140 mikrónov. Najdlhšia hladká myo-
cytocyty dosahujú až 500 mikrónov v stene maternice. Priemer myocytov je v rozsahu od 2 do 20 mikrónov. V závislosti od povahy intracelulárnych biosyntetických procesov sa rozlišujú kontraktilné a sekrečné myocyty. Prvý z nich sa špecializuje na funkciu kontrakcie, ale zároveň si zachováva sekrečnú činnosť.
Sekrečné myocyty sa vo svojej ultraštruktúre podobajú fibroblastom, ale obsahujú vo svojich cytoplazmatických zväzkoch tenké myofily umiestnené na periférii bunky. Golgiho komplex, granulované endoplazmatické retikulum, veľa mitochondrií, glykogénových granúl, voľné ribozómy a polysómy sú v cytoplazme dobre vyvinuté. Podľa stupňa zrelosti sú tieto bunky klasifikované ako zle diferencované. Aktínové vlákna v cytoplazme tvoria trojrozmernú sieť, ktorá je predĺžená hlavne pozdĺžne. Konce nekonečných vlákien sú navzájom spojené a k plazmolemme pomocou špeciálnych zosieťovacích proteínov. Tieto oblasti sú jasne viditeľné v elektrónových mikrografoch ako husté telá. Myozínové monoméry sa nachádzajú vedľa aktínových vlákien. Plazmoléma tvorí intrúzie - jaskyne, v ktorých sú koncentrované ióny vápnika. Signál kontrakcie zvyčajne prichádza cez nervové vlákna. Mediátor, ktorý sa odlišuje od ich termínov, mení priepustnosť plazmolémie. Uvoľňujú sa vápenaté ióny, čo spôsobuje polymerizáciu myozínu, ako aj interakciu myozínu s aktínom.
Medzi myozi dochádza k stiahnutiu aktínových myofilických vlákien
Obr. 9.12.Štruktúra hladkého myocytu (schéma):
a v- pri relaxácii; b. d- pri najväčšom znížení; g- s neúplným znížením; v-d- zväčšené obrázky orámovaných častí na fragmentoch aa b. 1 - plazmoléma; 2 - husté telá; 3 - jadro; 4 - endoplazma; 5 - kontraktilné komplexy; 6 - mitochondrie; 7 - suterénna membrána; 8 - aktínové (tenké) vlákna; 9 - myozínové (silné) myofily
Obr. 9.13.Ultraštruktúra diferenciačného hladkého myocytu v stene priedušiek:
1 - jadro; 2 - cytoplazma s myofilmi; 3 - Golgiho komplex, nárast o 35 000 (liek A. L. Zashikhina)
nové husté škvrny sa spoja, sila sa prenesie na plazmatém a celá bunka sa skráti (pozri obr. 9.12). Keď signály z nervového systému prestanú, vápnikové ióny sa pohybujú z cytoplazmy do jaskyne a tubuly endoplazmatického retikula, myozín sa depolymerizuje a myofibrily sa rozpadajú. Kontrakcia sa zastaví. Aktinomyozínové komplexy teda existujú v hladkých myocytoch iba počas obdobia kontrakcie v prítomnosti voľných iónov vápnika v cytoplazme.
Myocyty sú obklopené bazálnou membránou. V niektorých oblastiach sa v ňom vytvárajú „okná“, takže sa zbližuje plazma- léma susedných myocytov. Vytvárajú sa tu prípady a medzi bunkami vznikajú nielen mechanické, ale aj metabolické väzby. Na vrchnej časti „krytov“ z bazálnej membrány medzi myocytmi sú elastické a retikulárne vlákna, ktoré kombinujú bunky do jedného tkanivového komplexu. Hladké myocyty syntetizujú proteoglykány, glykoproteíny, prokolagén, pro-elastín, z ktorých sa tvoria kolagén a elastické vlákna a amorfná zložka medzibunkovej matrice. Interakcia myocytov sa vykonáva pomocou cytoplazmatických mostíkov, vzájomných výčnelkov, nexúz, desmozómov, rezov membránových kontaktov povrchov myocytov.
Regenerácie.Tkanivo hladkého svalstva viscerálnych a vaskulárnych druhov má významnú citlivosť na účinky extrémnych faktorov. V aktivovaných myocytoch sa zvyšuje úroveň biosyntetických procesov, ktorých morfologická expresia je syntéza kontraktilných proteínov, zväčšenie a hyperchromatóza jadra, hypertrofia jadra, zvýšenie jadrového cytoplazmatického pomeru, zvýšenie počtu voľných ribozómov a polysomov a

Obr. 9.14.Štruktúra tkaniva hladkého svalstva (objemová schéma) (podľa R. V. Krstika, so zmenami):
1 - fúzne hladké myocyty; 2 - cytoplazma myocytov; 3 - jadrá myocytov; 4 - plazmoléma; 5 - suterénna membrána; 6 - povrchové pinocytotické vezikuly; 7 - medzibunkové spojenia; 8 - nervové zakončenie; 9 - kolagénové vlákna; 10 - mikrovlákna
enzýmov, aeróbna a anaeróbna fosforylácia, membránový transport. Bunková regenerácia sa uskutočňuje tak v dôsledku diferencovaných buniek so schopnosťou vstúpiť do mitotického cyklu, ako aj v dôsledku aktivácie kambodžských prvkov (malé objemové myocyty). Pri pôsobení mnohých škodlivých faktorov je zaznamenaná fenotypová transformácia kontraktilných myocytov na sekrečné. Táto transformácia sa často pozoruje pri poškodení vaskulárnej intimy, tvorbe intimálnej hyperplázie s rozvojom aterosklerózy.
Obr. 9.15.Ultraštruktúra myopigmentocytov (liek N. N. Sarbaeva): 1 - jadro; 2 - myofilamenty, nárast o 6000
9.3.2. Mezenchymálne svalové tkanivo v orgánoch
Myocyty sú kombinované do zväzkov, medzi ktorými sú tenké vrstvy spojivového tkaniva. Do týchto vrstiev sú tkané retikulárne a elastické vlákna obklopujúce myocyty. Cez medzivrstvy prechádzajú krvné cievy a nervové vlákna. Konce posledne uvedených nekončia priamo na myocytoch, ale medzi nimi. Preto sa po príchode nervového impulzu mediátor šíri difúzne a vzrušuje mnoho buniek naraz. Hladké svalové tkanivo mezenchymálneho pôvodu je zastúpené hlavne v stenách krvných ciev av mnohých vnútorných dutých orgánoch.
Hladké svalové tkanivo v konkrétnych orgánoch má nerovnaké funkčné vlastnosti. Je to spôsobené skutočnosťou, že na povrchu orgánov existujú rôzne receptory pre špecifické biologicky aktívne látky. Preto pri mnohých liekoch nie je ich reakcia rovnaká. Je možné, že rôzne funkčné vlastnosti tkanív sú spojené so špecifickou molekulárnou organizáciou aktínových vlákien.
9.3.3. Svalové tkanivo nervového pôvodu
Svalové tkanivo dúhovky a ciliárneho tela je štvrtým typom kontraktilného tkaniva. Myocyty tohto tkaniva sa vyvíjajú z buniek nervového základu ako súčasť vnútornej steny optického pohárika. V rade
Obr. 9.16.Myoepiteliálne bunky v konečnej časti slinnej žľazy (schéma podľa G. S. Katinas):
a- prierez; b- pohľad z povrchu. 1 - jadrá myoepiteliálnych buniek; 2 - procesy myoepiteliálnych buniek; 3 - jadrá sekrečných epitelových buniek; 4 - suterénna membrána
prvky stavcov svalu dúhovky vykazujú rôznorodú divergentnú diferenciáciu. Teda myoneurálne tkanivo plazov a vtákov je predstavované pruhovanými viacjadrovými vláknami, ktoré sú veľmi podobné kostrovému svalstvu. U cicavcov a ľudí je hlavnou štrukturálnou a funkčnou jednotkou svalov dúhovky hladký mononukleárny myocyt alebo myopigmentocyt. Tieto majú pigmentované teleso obsahujúce jedno jadro, odobraté mimo vretenovitej kontraktívnej časti (obr. 9.15).
Cytoplazma buniek obsahuje veľké množstvo granúl mitochondrií a pigmentov, ktoré majú podobnú veľkosť a tvar ako granule pigmentového epitelu. Myofilamenty v myopigmentocytoch sa delia na tenké (7 nm) a silné (1,5 nm), svojou veľkosťou a umiestnením sa podobajú myofilamentom hladkých myocytov. Každý myopigmentocyt je obklopený bazálnou membránou. V blízkosti cytoplazmatických procesov myocytov sa nachádzajú nervové vlákna bez myelínu. V závislosti od smeru procesov (kolmých alebo rovnobežných s okrajom zornice) tvoria myocyty dva svaly - zúženie a zväčšenie zrenice.
Regenerácie.Niekoľko štúdií preukázalo nízku regeneračnú aktivitu po poškodení alebo neprítomnosti.
9.3.4. Epidermálne svalové bunky
Myoepiteliálne bunky sa vyvíjajú z epidermálneho zárodku. Nachádzajú sa v potných, mliečnych, slinných a slzných žľazách a majú spoločné prekurzory so svojimi sekrečnými bunkami. Myoepiteliální
bunky priamo susedia s vlastným epitelom a majú s nimi spoločnú bazálnu membránu. Počas regenerácie sa tieto a ďalšie bunky tiež obnovia z bežných nízko diferencovaných prekurzorov. Väčšina myoepiteliálnych buniek má tvar hviezdy. Tieto bunky sa často nazývajú kôšovité: ich procesy pokrývajú koncové úseky a malé kanáliky žliaz (obr. 9.16). Jadro a organely všeobecného významu sa nachádzajú v tele bunky a v procesoch sa nachádza kontraktilný aparát organizovaný ako v bunkách svalového tkaniva mezenchymálneho typu.
Bezpečnostné otázky
1. Genetická klasifikácia svalového tkaniva. Štrukturálne a funkčné jednotky rôznych typov svalového tkaniva.
2. Pružené tkanivo kostrového svalstva: vývoj, štruktúra, morfologický základ svalovej kontrakcie. Regenerácie.
3. Prúžkové srdcové svalové tkanivo: vývoj, špecifická štruktúra rôznych typov kardiomyocytov, regenerácia.
4. Druhy hladkých myocytov: zdroje vývoja, topografia v tele, regenerácia.
Histológia, embryológia, cytologia: učebnica / Yu. I. Afanasyev, N. A. Yurina, E. F. Kotovsky a ďalší. - 6. vydanie, Rev. a pridať. - 2012. - 800 s. : chorý.
Svaly ľudského tela sú tvorené hlavne svalovým tkanivom, ktoré sa skladá zo svalových buniek. Rozlišujte medzi hladkým a priečne pruhovaným svalovým tkanivom. Hladké svalové tkanivo - buduje svalstvo, ktoré je súčasťou niektorých vnútorných orgánov, a - žliabkovanýtvorí kostrový sval. Spoločnou vlastnosťou svalového tkaniva je jeho vzrušivosť, vodivosť a vtažitelnost.
Prúžkové svalové tkanivo sa odlišuje od hladkej, vyššej vzrušivosti, vodivosti a kontraktility. Bunky priečne pruhovaného svalu majú veľmi malý priemer a veľkú dĺžku (až 10 až 12 cm). V tejto súvislosti sa nazývajú vlákna.
Rovnako ako iné bunky, aj svalové bunky sa nazývajú protoplazmy sarcoplasm(z gréckeho. Sarcos - mäso). Nazýva sa membrána svalových buniek sarkolemou, Vo svalovom vlákne sú početné jadrá a ďalšie zložky buniek.
Zloženie svalových vlákien obsahuje veľké množstvo ešte tenších vlákien - myofibrils, ktoré zase pozostávajú z najtenších vlákien - protofibríl. protofibrilám - je to kontraktívny aparát svalovej bunky, jedná sa o špeciálne kontraktívne proteíny - myosin a aktínu, Mechanizmus svalových kontrakcií je komplexný proces fyzikálnych a chemických transformácií vo svalovom vlákne s povinnou účasťou kontraktilného aparátu. Tento mechanizmus je vyvolaný nervovým impulzom a energiu pre redukčný proces dodáva kyselina adenozíntrifosforečná (ATP). Z tohto hľadiska je znakom štruktúry svalových vlákien aj veľké množstvo mitochondrií, ktoré dodávajú svalovému vláknu potrebnú energiu. Relaxácia svalových vlákien je podľa mnohých vedcov uskutočňovaná pasívne v dôsledku elasticity sarkolemmy a intramuskulárneho spojivového tkaniva.
Štruktúra, tvar a klasifikácia kostrových svalov
Anatomická jednotka najaktívnejšej časti ľudského svalového systému - kostrové alebo priečne pruhované svaly - je kostrové svalstvo. Kostrový sval - Ide o orgán tvorený priečne pruhovaným svalovým tkanivom, ktoré obsahuje aj spojivové tkanivo, nervy a krvné cievy.
Každý sval je obklopený akýmsi „prípadom“ spojivového tkaniva - fascia.
v vonkajšia štruktúra svaly rozlišujú medzi šľachovou hlavou zodpovedajúcou začiatku svalu, bruchom svalu alebo telom tvoreným svalovými vláknami a koncom svalu šľachy alebo chvostom, ktorým sa sval pripája k inej kosti. Chvost svalu je spravidla pohyblivým bodom pripojenia a začiatok je pevný. V procese pohybu sa ich funkcie môžu meniť: body pohybu sa stanú nehybnými a naopak.
Tvar svalov je veľmi rôznorodý a vo veľkej miere závisí od funkčného účelu svalu. Rozlišujte medzi dlhými, krátkymi, širokými, okrúhlymi a ostatnými svalmi. dlho svaly sú umiestnené na končatinách krátky - tam, kde je pohyb malý (napríklad medzi stavcami). Široké svaly sa nachádzajú hlavne na tele (svaly brucha, chrbta, hrudníka). Kruhové svaly sú umiestnené okolo otvorov tela a pri sťahovaní ich sťahujú. Tieto svaly sa nazývajú zvierača, Ak má sval jednu hlavu, nazýva sa to jednoduché, ak je dva alebo viac - zložité (napríklad biceps, triceps a quadriceps). Dôležitým morfologickým znakom je umiestnenie svalových vlákien. Existuje paralelné, šikmé, priečne a kruhové usporiadanie vlákien (v zvieračoch). ak pri šikmom usporiadaní svalových vlákien priľnú na šľachách iba na jednej strane, svaly sa nazývajú semipenniformak na oboch stranách - dvuperistymi.
Funkčne je možné svaly rozdeliť na flexory a extensor, rotátory vonku a rotátory vo vnútri, svaly aduktora a prieduchy, Svaly sú tiež vylučované - synergenty a svaly - antagonisti, Prvý z nich tvorí skupinu svalov, priateľský pri každom pohybe, jeho kontrakcia spôsobuje opačné pohyby.
Hlavné svalové skupiny
Kostrové svaly dospelého človeka tvoria asi 40% jeho telesnej hmotnosti. U novorodencov a malých detí tvoria svaly maximálne 20 - 25% telesnej hmotnosti av starobe dochádza k postupnému znižovaniu svalovej hmoty na 25 - 30% telesnej hmotnosti. Celkovo má ľudské telo viac ako 600 svalov.
Podľa polohy, t. na topografickom základe sa rozlišujú svaly chrbta, hrudníka, brucha, hlavy, krku, horných a dolných končatín.
Zadné svaly sú rozdelené na povrch a hlboký, K povrchové svaly chrbty zahŕňajú: lichobežníkové svaly, latissimus dorsi, kosoštvorcové svaly (veľké a menšie), zubné svaly (horné a dolné).
Tieto svaly sa podieľajú na pohyboch hornej končatiny a na zmene objemu. hrudník (akt dýchania).
Hlboké svaly chrbát predstavuje množstvo svalov umiestnených pozdĺž chrbtice. Priťahujú sa najmä k stavcom stavcov a podieľajú sa na pohybe chrbtice späť a zboku.
Svaly hrudníka sa delia na svaly patriace do ramenného pletenca a horná časť končatiny (hlavné a menšie svaly pectoralis, svaly subclaviánu a predného dentátu) , a vlastné svaly hrudníka (vonkajšie vnútorné medzirebrové svaly). Dajú sa do pohybu a posilňujú horné končatiny na tele a podieľajú sa na fungovaní dýchania. Membrána tiež patrí do dýchacích svalov (oddeľuje hrudnú dutinu od brušnej dutiny).
Brušné svaly sú reprezentované vonkajšími a vnútornými šikmými, priečnymi a rektálnymi brušnými svalmi, ako aj štvorcovým svalstvom dolnej časti chrbta.
Svaly rectus abdominis sa podieľajú na prednom ohybe tela a šikmé svaly poskytujú sklon chrbtice do strán a jej zákruty spolu s hrudníkom doprava a doľava. Brušné svaly sa nezúčastňujú iba na pohyboch trupu a hrudníka. Tvoria stenu brušná dutina a vďaka svojmu tónu udržiavajú svoje vnútorné orgány v normálnej polohe. Ich znižovaním zvyšujú vnútrobrušný tlak a vytvárajú tzv abs, Brušné svaly uľahčujú pohyby čriev (stolice), močenie a vylučovanie plodu u žien počas pôrodu. Kontrakcia brušných svalov tiež prispieva k pohybu krvi v žilovom systéme ak uskutočňovaniu respiračných pohybov, ale tiež k tvorbe prietrže. Pri hernii dochádza k vyčnievaniu vnútorných orgánov (črevá, žalúdok atď.) Z brušnej dutiny pod kožou.
Medzi svaly hlavy sa rozlišujú žuvacie svaly a svaly tváre. K žuvacie svaly uplatňuje dočasné, žuvanie, pterygoid. Kontrakcia týchto svalov spôsobuje zložité žuvacie pohyby. dolná čeľusť, Mimické svaly sa líšia od ostatných svalov ľudského tela tým, že sú pripojené k pokožke tváre jedným alebo niekedy dvoma koncami. Kontrakcia týchto svalov je posunutie kože, tvorba záhybov a vrások. Toto definuje výrazy tváre, t. jeden alebo iný výraz tváre. Prejav komplexných pocitov (emócií) - radosť, hanba, pohŕdanie, smútok a bolesť atď. - Je určený početnými kombináciami kontrakcií svalov tváre, ktoré sú podriadené impulzom smerujúcim k nim z mozgovej kôry pozdĺž tvárového nervu. Kruhové svaly oka a úst sú tiež napodobňujúcimi svaly. Svaly krku nakláňajú hlavu, nakláňajú ju a otáčajú sa. Svaly spojené s hyoidnou kosťou počas kontrakcie menia polohu jazyka a hrtanu pri prehĺtaní a vydávaní zvukov.
Svaly hornej končatiny sa delia na svaly ramenný opasok a svaly voľnej hornej končatiny. Svaly ramenného pletenca obklopujú ramenný kĺb a poskytujú v ňom početné pohyby. Medzi nimi je najdôležitejší deltový sval. Pri sťahovaní tento sval ohýba ruku ramenný kĺb a vezme ruku do vodorovnej polohy.
Medzi svaly voľnej hornej končatiny patria svaly ramena, predlaktia a ruky.
V oblasti ramien vpredu je svalová skupina - flexoryspäť - extensor, Medzi svaly prednej skupiny je najdôležitejší sval bicepsu na ramene (biceps) a chrbát, sval tricepsu na ramene.
Svaly predlaktia sú tiež predstavované na prednej ploche ohýbačmi a na zadnej strane extenzormi. Svaly rúk sa nachádzajú iba na povrchu dlane. Medzi nimi vyniká - dlhá dlane svaly a ohnutie prstov. Vzhľadom na prítomnosť vlastného svalového systému, najmä prstov palec, získať väčšiu mobilitu a sú schopné rôznych pohybov vrátane maximálneho ohybu a roztiahnutia, čo je pri práci mimoriadne dôležité.
Svaly nachádzajúce sa v oblasti pásu dolnej končatiny posúvajú nohu dovnútra bedrový kĺba rovnako s pevnou končatinou ohýbajú chrbticu v bedrovej oblasti. Predná svalová skupina obsahuje jeden hlavný sval - iliopsoas. Medzi zadnou vonkajšou skupinou svalov panvového pletenca - veľké, stredné a malé gluteálne svaly.
Dolné končatiny majú masívnejšiu kostru ako horné; ich svaly majú veľkú silu, ale zároveň menšiu rozmanitosť a obmedzený rozsah pohybu.
Na stehne vpredu je najdlhší prispôsobený sval v ľudskom tele (do 50 cm). Ohýba nohu v bedrových a kolenných kĺboch.
Štvorhlavý svaly femoris leží hlbšie ako svaly na mieru a obklopujú stehennú časť takmer zo všetkých strán.
Hlavnou funkciou tohto svalu je predĺženie kolenného kĺbu. Keď stojí, štvorhlavý sval nedáva kolenný kĺb ohnúť.
Na chodidle sa rozlišujú chrbtové a plantárne svaly. Tieto svaly sa podieľajú na ohýbaní a predlžovaní prstov na nohách, ako aj na ich zbližovaní a šľachtení.