Сколько крестцовых позвонков у ящерицы. Опорно-двигательный аппарат пресмыкающихся общая характеристика скелета
ТЕМА 14. СКЕЛЕТ ПРЕСМЫКАЮЩЕГОСЯ
СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ ОБЪЕКТА
Подтип Позвоночные, Vertebrata
Класс Пресмыкающиеся, Reptilia
1. Отряд Чешуйчатые, Squamata
Представитель - Серый варан, Varanus griseus Daud.
2. Отряд Крокодилы, Crocodilia
Представитель - Миссисипский аллигатор, Alligator mississippiensts Daud.
3. Отряд Черепахи, Chelonia (Testudines)
Представитель - Болотная черепаха, Etnys orbicular is L.
МАТЕРИАЛ И ОБОРУДОВАНИЕ
Смонтированные скелеты варана, змеи, черепахи - по одному на группу студентов; черепа крокодилов (или муляжи) - 1-2 на группу. На одного-двух студентов необходимы:
1. Разборный скелет варана.
2. Череп черепахи.
3. Препарировальные иглы - 2.
ЗАДАНИЕ
Рассмотреть скелет варана в целом. Найти его основные части: череп, позвоночник, скелет парных конечностей и их поясов. Рассмотреть особенности строения скелета змеи и черепахи. Рассмотреть детали строения скелета варана и черепов варана, крокодила и черепахи. Сравнить строение черепа крокодила, варана, змеи и черепахи.
Сделать следующие рисунки:
1. Череп варана сбоку.
2. Череп варана снизу.
4. Передняя конечность варана.
6. Задняя конечность варана.
7. Череп крокодила сверху.
8. Череп крокодила снизу.
9. Череп черепахи сбоку.
ОПИСАНИЕ СКЕЛЕТА
Осевой скелет. Дифференцировка осевого скелета, или позвоночника, на отделы выражена у пресмыкающихся значительно отчетливее, чем у земноводных. Шейный отдел (pars cervicalis) всегда составлен несколькими позвонками, из которых два передних имеют особое устройство. Первый шейный позвонок называется атлас или атлант (atlas; рис. 81, А). Он лишен тела позвонка и имеет форму разделенного на две части кольца. На нижней передней поверхности этого позвонка имеется сочленовная впадина, подвижно соединяющаяся с мыщелком черепа (см. ниже). Второй шейный позвонок - эпистрофей (epistrophaeus; рис. 81, Б), имеет спереди крупный зубовидный отросток (processus odontoideus, рис. 81, 1), который представляет собой тело первого шейного позвонка, сросшееся с эпистрофеем. Зубовидный отросток свободно входит в нижнее отверстие атласа. Такое строение первых шейных позвонков обеспечивает большую подвижность головы. Остальные шейные позвонки имеют обычное устройство (см. ниже); многие из них несут короткие шейные ребра.
Рис. 81. Позвонки варана
А - атлант; Б - эпистрофей; В - грудной позвонок; Г - продольный разрез грудного позвонка:
1 - зубовидный отросток эпистрофея, 2 - тело позвонка, 3 - верхняя дуга, 4 - остистый отросток, 5 - канал для спинного мозга, 6 - передний сочленовный отросток, 7 - задний сочленовный отросток
Грудной и поясничный отделы различаются не вполне отчетливо и обычно рассматриваются как единый отдел (pars thoracolumbalis). Собственно грудным отделом считается та часть позвоночника, в которой отходящие от позвонков ребра нижним концом причленяются к грудине. Позвонки поясничного отдела несут ребра, не доходящие до грудины. Тела позвонков (рис. 81, 2) спереди вогнутые, а сзади выпуклые; такие позвонки носят название процельных. Над телом позвонка поднимаются верхние дуги (arcus neuralis; рис. 81, 3), заканчивающиеся остистым отростком (processusspinosus; рис. 81, 4). В канале, образованном верхними дугами, располагается спинной мозг.
От переднего и заднего отделов основания верхних дуг отходят соответственно передние (processus articularis anterior; рис. 81, 6) и задние (processus articularis posterior; рис. 81, 7) сочленовные отростки. Эти парные отростки соединяются с сочленовными отростками соседних позвонков и способствуют большей прочности позвоночника при изгибах. По бокам на теле позвонка (вблизи основания верхних дуг) имеются небольшие углубления, к которым причленяются ребра.
Крестцовый отдел (pars sacralis) состоит из двух позвонков, для которых характерны мощно развитые поперечные отростки (processus transversus); к ним присоединяются кости таза. Хвостовой отдел (pars caudalis) представлен многочисленными позвонками, постепенно уменьшающимися в размерах.
Такое строение позвоночника типично для класса пресмыкающихся, но в некоторых группах оно претерпевает вторичные изменения. В частности, у змей в связи с редукцией парных конечностей и возникновением иного типа передвижения - переползания на брюхе путем изгибов туловища - позвоночник отчетливо делится лишь на туловищный и хвостовой отделы. Все туловищные позвонки имеют подвижные ребра, нижние концы которых свободны (грудина у змей отсутствует) и упираются в брюшные роговые щитки.

Рис. 82. Скелет болотной черепахи
А - карапакс; Б - пластрон:
1 - туловищный отдел позвоночного столба, 2 - реберные пластинки, 3 - краевые пластинки, 4 - коракоид, 5 - лопатка, 6 - подвздошная кость, 7 - лобковая кость, 8 - седалищная кость
У черепах осевой скелет принимает участие в образовании костной основы их панциря. Верхний щит панциря - карапакс (caparax; рис. 82, А) - составлен несколькими рядами костных пластинок. Средний (непарный) ряд этих пластинок образован срастанием расширенных и уплощенных остистых и поперечных отростков туловищных позвонков с кожными костями; по бокам от среднего ряда лежат парные ряды костных пластинок, сросшихся с расширенными ребрами. Край карапакса образуют костные пластинки покровного происхождения. Таким образом, туловищный отдел позвоночника черепах неподвижен и прочно сращен со спинным щитом панциря. Шейный же и хвостовой отделы позвоночника - подвижны. При этом передние шейные позвонки опистоцельные (тело позвонка спереди выпуклое, сзади вогнутое), задние - продельные, а между этими двумя группами располагается один позвонок, тело которого имеет выпуклую поверхность и спереди и сзади.
Череп. По сравнению с земноводными череп пресмыкающихся характеризуется значительно более полным окостенением. Некоторое количество хряща сохраняется лишь в обонятельной капсуле и в слуховой области. Осевой и висцеральный отделы черепа эмбрионально закладываются отдельно, но у взрослых животных представляют собой единое образование. В состав черепа входят как хрящевые (замещающие, или первичные), так и многочисленные кожные (покровные, или вторичные) кости. В качестве основного объекта для изучения удобно использовать череп крупной ящерицы - варана.

Рис. 83. Череп варана
А - сбоку; Б - снизу; В - сверху; Г - сзади:
1 - основная затылочная кость, 2 - боковая затылочная кость, 3 - верхняя затылочная кость, 4 - большое затылочное отверстие, 5 - затылочный мыщелок, 6 - переднеушная кость, 7 - основная клиновидная кость, 8 - сошник, 9 - теменная кость, 10 - лобная кость, 11 - носовая кость, 12 - предлобная кость, 13 - предглазничная кость, 14 - слезная кость, 15 - верхняя височная яма, 16 - заднелобная кость, 17 - чешуйчатая кость, 18 - предчелюстная кость, 19 - верхнечелюстная кость, 20 - скуловая кость, 21 - разрыв нижней височной дуги благодаря редукции квадратно-скуловой кости, 22 - квадратная кость, 23 - крыловидная кость, 24 - нёбная кость, 25 - верхнекрыловидная кость, 26 - поперечная кость, 27 - надугловая кость, 28 - зубная кость, 29 - угловая кость, 30 - сочленовная кость, 31 - венечная кость
Осевой череп. В затылочном отделе черепа имеются все четыре затылочные кости: основная затылочная (basioccipitale, рис. 83, 1), две боковые затылочные (occipitale laterale; рис. 83, 2) и верхняя затылочная (supraoccipitale; рис. 83, 3). Эти первичные по происхождению кости окружают большое затылочное отверстие (foramen occipitale magnum; рис. 83, 4). Нижняя и боковые затылочные кости совместно образуют единственный (в отличие от земноводных) затылочный мыщелок (condylus occipitalis; рис. 83, 5), подвижно сочленяющийся с первым шейным позвонком - атласом. Сочленение головы с шеей при помощи только одного мыщелка в сочетании с уже рассмотренными особенностями строения двух первых шейных позвонков придает голове рептилий значительную подвижность.
В слуховом отделе из хрящевых костей сохраняет самостоятельность только парная переднеушная кость (prooticum; рис. 83, 6), тогда как верхнеушные срастаются с верхней затылочной костью, а задне-ушные - с боковыми затылочными.
Межглазничная перегородка у пресмыкающихся тонкая, перепончатая, и лишь у крокодилов и ящериц в ней имеются отдельные небольшие окостенения, видимо, соответствующие глазо-клиновидным костям. Обонятельная капсула окостенений не имеет.
В основании черепа впереди от основной затылочной кости располагается довольно крупная покровная основная клиновидная кость (basisphenoideum; рис. 83, 7). Ее передний узкий отросток гомологичен парасфеноиду (parasphenoideum), который у рептилий заметно редуцирован. В передней части дна черепа под обонятельным отделом расположен парный сошник (vomer; рис. 83, 8), также имеющий покровное происхождение.
Крыша черепа представлена многочисленными покровными костями, некоторые из которых опускаются книзу и прикрывают череп с боков. К ним относятся теменные (parietale; рис. 83, 9), лобные (frontale; рис. 83, 10) и носовые (nasale; рис. 83, 11) кости. Впереди лобных костей обычно располагаются парные предлобные (praefrontale; рис. 83, 12) и предглазничные (praeorbitale; рис. 83, 13) кости, а под ними в передней стенке глазницы - прободенные узким каналом парные слезные (lacrimale; рис. 83, 14).
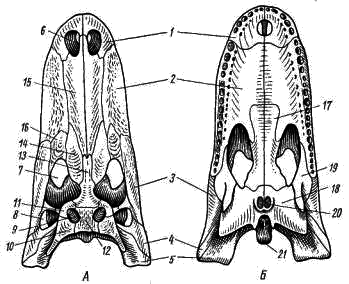
Рис. 84. Череп крокодила (миссисипский аллигатор)
.
А - сверху; Б - снизу:
1 - предчелюстная кость, 2 - верхнечелюстная кость, 3 - скуловая кость, 4 - квадратно-скуловая кость, 5 - квадратная кость, 6 - наружная ноздря, 7 - глазница, 8 -боковая височная яма, 9 - верхняя височная яма, 10 - чешуйчатая кость, 11 - заднелобная (заглазничная) кость, 12 - теменная кость, 13 - лобная кость, 14 - предлобная кость, 15 - носовая кость, 16 - слезная кость, 17 - нёбная кость, 18 - крыловидная кость, 19 - поперечная кость, 20 - хоаны (внутренние отверстия ноздрей), 21 - затылочный мыщелок
Из остальных покровных костей осевого черепа особый интерес представляют кости, принимающие участие в образовании так называемых височных дуг. У крокодила в крыше черепа кнаружи от теменной кости (рис. 84, 12) с каждой стороны имеется отверстие - верхняя височная яма (рис. 84, 9). По наружному краю верхняя височная яма ограничена заднелобной или заглазничной (postfrontale-postorbitale, рис. 84, 11) и чешуйчатой (squamosum; рис. 84, 10) костями. Эти две кости вместе составляют верхнюю височную дугу. Сбоку черепа позади глазницы располагаются боковые височные ямы (рис. 84, 8), ограниченные снаружи нижними височными дугами. Каждая нижняя височная дуга составлена двумя костями: скуловой (jugale; рис. 84, 3) и квадратно-скуловой (quadrato-jugale; рис. 84, 4). Нижняя височная дуга соединяется с верхней челюстью: скуловая кость прирастает к верхнечелюстной, а квадратно- скуловая - к квадратной. Такой тип черепа, как у крокодила - с двумя височными ямами и двумя височными дугами, носит название диапсидного (двудужного).
У варана верхняя височная яма (рис. 83, 15) ограничена полной верхней височной дугой (postfrontale - squamosum; рис. 83, 16-17), В составе же нижней височной дуги редуцировалась квадратно-скуловая кость и сохранилась только скуловая (рис. 83, 20); боковые височные ямы вследствие этого незамкнуты снаружи и остаются открытыми. Поэтому череп варана может рассматриваться как череп диапсидного типа, но с редуцированной нижней дугой. У некоторых других ящериц частично редуцируется и верхняя височная дуга, а у змей (см. рис. 86) обе височные дуги редуцированы (заднелобная и чешуйчатая кости не соединяются друг с другом; обе височные ямы снаружи остаются открытыми).
Таким образом, змеи и ящерицы (отряд чешуйчатых, Squamata) по строению черепа относятся к группе диапсидных (двудужных) пресмыкающихся, но характеризуются разной степенью редукции височных дуг.

Рис. 85. Череп болотной черепахи
:
1 - ложная височная яма, 2 - предчелюстная кость, 3 - верхнечелюстная кость, 4 - скуловая кость, 5 - квадратно-скуловая кость, 6 - квадратная кость, 7 - чешуйчатая кость, 8 - заднелобная кость, 9 - теменная кость, 10 - лобная кость, 11 - предлобная кость, 12 - верхняя затылочная кость
У черепахи обе височные ямы отсутствуют, и боковая стенка крыши черепа, отграничивающая снаружи большую полость - так называемую ложную височную яму (рис. 85, 1), образовавшуюся как выемка в затылочной части черепа, составлена плотно сросшимися костями: заднелобной (postfrontale; рис. 85, 8), чешуйчатой (squamosum; рис. 85, 7), скуловой (jugale; рис. 85, 4) и квадратно-скуловой (quadrato-jugale, рис. 85, 5). Такой тип черепа, лишенный истинных височных ям и ограничивающих их височных дуг, называется анапсидным (бездужным).
Висцеральный череп. У варана нёбно-квадратный хрящ окостеневает, образуя в заднем отделе квадратную кость (quadratum; рис. 83, 22), к нижнему концу которой причленяется нижняя челюсть; верхний конец квадратной кости подвижно сочленен с осевым черепом. Впереди квадратной кости расположена крыловидная кость (pterygoideum; рис. 83, 23), а перед ней - нёбная кость (palatinum; рис. 83, 24), соединяющаяся с верхнечелюстными костями и сошником. Все эти кости парные; из них только квадратные кости имеют хрящевое (первичное) происхождение.
От крыловидной кости вверх отходит верхнекрыловидная кость (epipterygoideum; рис. 83, 25). Эта парная кость, соединяющая крыловидную и теменную кости, гомологична вертикальному («восходящему») отростку нёбно-квадратного хряща и характерна из ныне живущих рептилий для ящериц и гаттерии. Кроме верхнекрыловидных от крыловидных костей отходят поперечные кости (transversum; рис. 83, 26), которые в передней своей части присоединяются к верхнечелюстным костям. Вторичная верхняя челюсть представлена предчелюстными (praemaxillare; рис. 83, 18) и верхнечелюстными (maxillare; рис. 83, 19) костями. Нижняя челюсть состоит из первичной сочленовной кости (articulare; рис. 83, 30) и покровных костей: зубной (dentale; рис. 83, 28), угловой (angulare; рис. 83, 29), надугловой (supraangulare; рис. 83, 27), венечной (coronare; рис. 83, 31) и, иногда, еще нескольких мелких косточек.
На предчелюстной, челюстной и зубной костях рептилий (кроме черепах) расположены простые конические, иногда слегка загнутые назад зубы, которые прирастают к краю соответствующей кости.
Подъязычная дуга, как и у земноводных, полностью утратила функцию подвеска. Верхний элемент подъязычной дуги (гиомандибуляре) входит в состав среднего уха в виде палочковидной слуховой косточки - стремечка (stapes seu columella), а остальная ее часть вместе с остатками передних жаберных дуг образует подъязычный аппарат.
Описанное строение висцерального черепа в общем типично для всех пресмыкающихся. Но в некоторых группах имеются отступления от этой схемы, связанные главным образом со спецификой биологии этих групп.

Рис. 86 Череп ядовитой змеи
:
1 - предчелюстная кость, 2 - верхнечелюстная кость. 3 - нёбная кость, 4 - крыловидная кость, 5 - поперечная кость, 6 - квадратная кость, 7 - чешуйчатая кость, 8 - заднелобная кость, 9 - ядовитый зуб, 10 - лобная кость, 11 - носовая кость, 12 - зубная кость. 13 - угловая кость, 14 - сочленовная кость
У змей очень подвижны не только квадратные (рис. 86, 6), но и соединенные с ними чешуйчатые (рис. 86, 7), а также крыловидные (рис. 86, 4) и нёбные (рис. 86, 3) кости. Две последние несут острые зубы. Поперечные кости (рис. 86, 5) у змей служат как бы рычагами, передающими движения крыловидных костей верхнечелюстным костям (рис. 86, 2), которые в свою очередь очень подвижны. Вся эта система подвижно сочлененных костей не только способствует чрезвычайно широкому раскрыванию рта, но и обеспечивает независимые движения правой и левой половин челюстного аппарата при проталкивании добычи в глотку с поочередным перехватыванием. Это позволяет змеям заглатывать относительно очень крупную (превышающую толщину туловища змеи) добычу. У ядовитых змей на верхнечелюстных костях расположены подвижно прикрепленные острые, загнутые назад ядовитые зубы (рис. 86, 9), имеющие внутренний канал или бороздку на передней поверхности, по которым при укусе в ранку стекает яд из расположенных у основания зуба ядовитых желез.
Череп крокодилов (рис. 84) характеризуется тем, что зубы не прирастают к краю зубной, предчелюстной и верхнечелюстной костей, как у других рептилий, а сидят в специальных углублениях (лунках, или альвеолах) этих костей - текодонтные зубы. Другая особенность висцерального черепа крокодилов - вторичное твердое нёбо, отделяющее ротовую полость от носоглоточного хода.
В образовании вторичного твердого нёба принимают участие нёбные отростки предчелюстных (рис. 84,7) и верхнечелюстных (рис. 84, 2) костей, а также нёбные (рис. 84, 17) и крыловидные (рис. 84, 18) кости. Благодаря образованию твердого нёба вторичные хоаны (рис. 84, 20) отнесены назад и расположены в крыловидных костях, над гортанью. Образование вторичного твердого нёба связано с характером образа жизни крокодилов: прямое соприкосновение гортани с хоанами открывает возможность бесперебойного дыхания при принятии пищи и когда крокодил отдыхает на мелководье, выставив из воды находящиеся на возвышениях ноздри, тогда как ротовая полость заполнена водой.
Парные конечности и их пояса. Плечевой пояс пресмыкающихся состоит из типичных костей: расположенной более дорзально лопатки (scapula; рис. 87, 1) и обращенного в вентральную сторону коракоида (coracoideum; рис. 87, 3). Обе эти кости принимают участие в образовании суставной ямки (рис. 87, 4) для прикрепления передней конечности. Дорзальнее лопатки располагается широкий уплощенный надлопаточный хрящ (cartilago suprascapularis; рис. 87, 2), а впереди коракоида - хрящевой прокоракоид (cartilago procoracoidea; рис. 87, 5). Имеется хорошо развитая грудина (sternum, рис. 87, 6), к которой причленяется несколько ребер (costa, рис. 87, 7). Таким образом, в отличие от земноводных у рептилий развивается грудная клетка и плечевой пояс имеет опору в осевом скелете. На вентральной стороне грудины располагается Т- образная покровная костъ - надгрудинник (episternum; рис. 87, 8), впереди нее - тоже покровные кости - ключицы (clavicula; рис. 87, 9). Наружные концы ключиц причленяются к лопаткам, а внутренние срастаются с ветвями надгрудинника. Ключицы и надгрудинник (отсутствующий у земноводных) увеличивают прочность соединения правой и левой частей плечевого пояса.

Рис. 87. Плечевой пояс варана (вид снизу)
:
1 - лопатка, 2 - надлопаточный хрящ, 3 - коракоид, 4 - суставная впадина для головки плеча, 5 - прокоракоидный хрящ, 6 - грудина, 7 - ребра, 8 - надгрудинник, 9 - ключица
У змей плечевой пояс полностью редуцирован, а у черепах ключицы и надгрудинник вошли в состав костей брюшного щита (plastron; см. рис. 82, Б) панциря, образовав соответственно передние парные и вклинившуюся между ними непарную костные пластинки.

Рис. 88. Тазовый пояс варана (вид снизу)
:
1 - подвздошная кость, 2 - лобковая кость, 3 - седалищная кость, 4 - вертлужная впадина (сочленовная ямка) для головки бедра, 5 - крестцовые позвонки
Тазовый пояс состоит из двух симметричных половин, соединенных по средней линии хрящом. Каждая половина составлена тремя костями; расположенной дорзально подвздошной (ilium; рис. 88, 1), находящимися на вентральной стороне лобковой (pubis; рис. 88, 2) и седалищной (ischium, рис. 88, 3). Все эти кости принимают участие в образовании сочленовной ямки (acetabulum; рис. 88, 4), к которой причленяется задняя конечность. Таз у рептилий закрытый: правая и левая лобковые и седалищные кости на брюшной стороне сращены между собой.

Рис. 89. Конечности варана
А - передняя; Б - задняя:
1 - плечевая кость, 2 - локтевая кость, 3 - лучевая кость, 4 - запястье, 5 - пясть, 6 - фаланги пальцев, 7 - интеркарпальный сустав, 8 - бедренная кость, 9 - большая берцовая кость, 10 - малая берцовая кость, 11 - коленная чашечка, 12 - предплюсна, 13 - интертарзальный сустав, 14 - плюсна
Конечности пресмыкающихся построены по типичной схеме конечностей наземных позвоночных. Проксимальный отдел передней конечности (рис. 89, А) представлен одной костью - плечевой (humerus; рис. 89, 1), далее следует предплечье, состоящее из двух костей - локтевой (ulna; рис. 89, 2) и лучевой (radius; рис. 89, 3). Запястье (carpus, рис. 89, 4) состоит из относительно мелких косточек, располагающихся обычно в два ряда; сбоку от них находится еще одна косточка - грушевидная, принимаемая за остаток шестого пальца. Пясть (metacarpus; рис. 89, 5) составлена пятью удлиненными косточками, к которым прикрепляются фаланги пяти пальцев (phalanges digitorum; рис. 89, 6).
Последние фаланги несут когти. Сустав, обеспечивающий подвижность кисти, у рептилий проходит не между костями предплечья и проксимальным рядом костей запястья (как у амфибий), а между проксимальным и дистальным рядами костей запястья. Такой сустав называется интеркарпальным (рис. 89, 7).
В задней конечности проксимальный элемент - бедро (femur; рис. 89, 8) сочленяется коленным суставом с голенью, состоящей из двух берцовых костей - большой (tibia; рис. 89, 9) и малой (fibula; рис. 89, 10). Над передней поверхностью этого сустава располагается маленькая косточка - коленная чашечка (patella; рис. 89, 11). В предплюсне (tarsus; рис. 89, 12) проксимальный ряд косточек срастается или почти неподвижно соединяется с костями голени, а косточки дистального ряда также тесно связаны и частично сращены с плюсневыми костями. Благодаря этому суставная поверхность здесь расположена не между голенью и стопой, а между проксимальным и дистальным рядами косточек предплюсны. Такой сустав характерен для пресмыкающихся и носит название интертарзального сустава (рис. 89, 13). Плюсна (metatarsus; рис. 89, 14) состоит из пяти удлиненных костей, к которым прикреплены фаланги пяти пальцев (phalanges digitorum; рис. 89, 6). Концевые фаланги несут когти.
ЗАКЛЮЧЕНИЕ
Пресмыкающиеся (Reptilia) - первый класс группы настоящих наземных позвоночных (Amniota), представители которого обладают комплексом приспособлений к обитанию на суше вне всякой связи с водной средой.
Выше было сказано, что тесная связь предков рептилий - земноводных - с водоемами (или по крайней мере с местами повышенной влажности) была обусловлена в первую очередь двумя биологическими особенностями: водным типом размножения и двойственностью органов дыхания (легкие и кожа). Эти особенности отражаются в свою очередь на характере водного обмена: обитая во влажной среде, амфибии не обладают морфофизиологическими механизмами уменьшения поте ри влаги организмом. Полное освоение суши немыслимо без коренных преобразований типа размножения, дыхания и водного обмена.
Осуществление этих преобразований характеризует класс пресмыкающихся.
Наиболее специфические особенности этого класса (и других амниот) как первично наземных позвоночных связаны с приспособлением к жизни в условиях относительно сухого атмосферного воздуха. Специфические черты строения рептилий в первую очередь связаны именно с приспособлением к более экономному расходованию влаги. Кожа пресмыкающихся сухая и лишена желез (кроме небольшого числа специализированных желез, выделяющих относительно густой секрет). Наружные слои эпидермиса представлены отмершими клетками, заполненными роговым веществом (кератогиалином).
Этот роговой слой кожи предохраняет организм от высыхания, а также в значительной степени улучшает защиту от химических и механических повреждений. Ниже рогового слоя расположен мальпигиев слой эпидермиса, представленный живыми клетками. По мере снашивания ороговевших клеток на смену им приходят клетки из нижних слоев, в протоплазме которых вначале появляются отдельные зернышки кератогиалина, заполняющие впоследствии всю клетку. Кроме общего ороговения верхнего слоя эпидермиса для рептилий характерно образование в коже различной формы роговых чешуи, покрывающих поверхность кожи. Чешуи осуществляют механическую защиту кожи и способствуют еще большему сокращению влагопотерь через покровы; у некоторых современных рептилий (например, у крокодилов) под роговой чешуей имеются костные пластинки.
Все эти преобразования, биологически выгодные с точки зрения поддержания более экономного водного баланса организма, оказались возможными прежде всего потому, что кожа рептилий не участвует в дыхании, а это связано с прогрессивным развитием дыхательной системы (см. ниже).
Важное значение в эволюции высших наземных позвоночных имеет преобразование выделительной системы. В отличие от ранее изученных классов пресмыкающиеся (как и другие Amniota) обладают тазовыми почками (metanephros). В отличие от туловищных почек (mesonephros) анамний выделительные канальцы тазовой почки полностью утратили связь с полостью тела; продукты обмена поступают внутрь выделительных канальцев только из сплетения кровеносных капилляров - мальпигиева клубочка. Он заключен в полом выросте концевой части канальца - боуменовой капсуле. Это приводит к тому, что в процессе выделения продуктов диссимиляции организм теряет меньше воды (не выводится полостная жидкость). Кровеносные сосуды, выходящие из мальпигиева клубочка, образуют вторую сеть капилляров вокруг стенки выделительного канальца, а затем впадают в более крупные сосуды венозной системы. Через эту вторую сеть капилляров происходит обратное всасывание в кровь воды, углеводов и некоторых солей, благодаря чему выделяемая так называемая «вторичная моча» становится более концентрированной и имеет иной состав, нежели «первичная моча», образующаяся в боуменовых капсулах.
Тазовая (метанефрическая) почка имеет свой мочеточник, не гомологичный ни мюллерову, ни вольфову каналам. Вольфов канал сохраняется только у самцов, где он через придаток семенника (epididymis) связан с половой железой и играет роль семяпровода. Мюллеров канал у самцов редуцирован. Самки же, напротив, сохраняют только мюллеровы каналы; они выполняют функцию яйцеводов и открываются в полость тела отверстиями - воронками яйцеводов. Вольфовы каналы у самок редуцированы.
Таким образом, главные особенности выделительной системы высших наземных позвоночных заключаются в утрате связи почек с полостью тела и в усилении системы обратного всасывания воды из первичной мочи в кровь. Эти свойства обеспечивают осуществление выделительной функции при минимальном расходовании воды, что для обитателей суши, как уже говорилось, имеет большое биологическое - значение. К этому можно добавить, что и среди наземных позвоночных животных интенсивность общей фильтрации и обратного всасывания воды различна у животных, обитающих в разных условиях. Так, среди рептилий крокодилы и черепахи (в основном пресноводные формы) имеют хорошо развитые клубочки и выделяют жидкую мочу, тогда как у ящериц и змей клубочки развиты заметно слабее, а моча выделяется твердая или полужидкая.
Весь комплекс рассмотренных особенностей, обеспечивающих более экономный водный баланс рептилий, лежит в основе окончательного отрыва высших наземных позвоночных от водной среды и широкого заселения ими разнообразных биотопов суши.
Наряду с этими особенностями для рептилий характерен ряд преобразований, имеющих общее прогрессивное значение в эволюции позвоночных.
Существенно изменилась система дыхания. Появилась трахея, длина которой зависит от длины шеи. Для легких пресмыкающихся характерна более сложная внутренняя структура, чем у земноводных: в них имеются внутренние перекладины, делящие их на отделы. От этих перекладин, в свою очередь, отходят перегородки, развитые в различной степени у разных групп. Между перегородками образуются внутрилегочные дыхательные пути, которые служат как бы продолжением бронхов и в ряде случаев делятся на внутрилегочные бронхи второго порядка. Внутренние стенки камер, образуемых всей этой системой перекладин, имеют ячеистую структуру, так что в результате легкие в целом приобретают сложную губчатую структуру. Такое устройство легких существенно увеличивает общую дыхательную поверхность, а соответственно и интенсивность дыхания. Благодаря этим приспособлениям потребность организма рептилий в кислороде полностью обеспечивается одним только легочным дыханием.
Крайне важно, что акт дыхания у пресмыкающихся осуществляется путем активного расширения и сжатия грудобрюшной полости под действием брюшной и реберной мускулатуры. Такой тип дыхания обеспечивает более интенсивную вентиляцию легких большим объемом воздуха; появление его связано с развитием подвижной грудной клетки.
Существенные прогрессивные преобразования происходят и в кровеносной системе пресмыкающихся (схема разделения потоков венозной и артериальной крови в сердце рептилий была дана при описании органов кровообращения).
Благодаря появлению неполной перегородки в желудочке и самостоятельному отхождению от разных его отделов главных артериальных стволов (что связано с редукцией артериального конуса) достигается достаточно полное разделение потоков артериальной и венозной крови. В момент систолы желудочка его внутренние полости на какое-то время оказываются изолированными; при этом в легочную артерию попадает венозная кровь, в правую дугу аорты - артериальная, а в левую дугу аорты - артериальная с примесью венозной. Поскольку обе сонные и подключичные артерии берут начало от правой дуги аорты, голова и передняя часть туловища снабжаются практически чистой артериальной кровью (так, у черепах в сонных артериях насыщение крови кислородом в половине случаев превышало 90%). Сосуды, отходящие ниже соединения дуг аорты, несут к органам задней половины тела кровь, содержащую определенную примесь венозной.
Горизонтальная перегородка, делящая желудочек сердца пресмыкающихся на две главные камеры, не гомологична межжелудочковой перегородке птиц и млекопитающих. Из всех групп пресмыкающихся только у крокодилов в желудочке появляется новая - вертикальная - перегородка, замещающая собой древнюю горизонтальную. Эта вертикальная перегородка полностью делит желудочек сердца крокодилов на две изолированные камеры: правую (венозную) и левую (артериальную). Таким образом, сердце крокодилов может быть названо четырех-камерным: в нем имеются два изолированных друг от друга предсердия и два также полностью разделенных желудочка. Однако наряду с полным разделением желудочков у крокодилов сохраняются обе дуги аорты, образующие характерный для всех рептилий перекрест: правая начинается в левом, а левая - в правом желудочке. В месте перекреста имеется так называемое паннициево отверстие, соединяющее полости обеих дуг аорты.
Сонные и подключичные артерии, как и у всех рептилий, отходят от правой дуги аорты. Левая дуга аорты, обогнув сердце, отдает сосуды к кишечнику и тонким протоком соединяется с правой дугой аорты, давая начало спинной артерии. По некоторым данным, в основании левой дуги аорты имеются клапаны и система хрящей, препятствующих поступлению в нее венозной крови при систоле желудочка.
Сохранение обеих дуг аорты и их связь в месте перекреста некоторые авторы объясняют экологическими особенностями крокодилов: во время пребывания под водой легочный круг кровообращения пропускает мало крови, так как дыхание приостановлено, и избыток крови из легочного круга кровообращения переходит в большой круг, поддерживая таким образом в его сосудах нормальное давление крови. Возможно, что такое же значение имеет и сохранение боталловых протоков (сосудов, соединяющих дуги аорты с легочными артериями) у черепах, ведущих водный образ жизни. Как и сонные протоки (соединение дуг аорты с сонными артериями) змей и ящериц, эти образования представляют собой рудименты артериальных дуг далеких предков рептилий, редуцировавшиеся при смене жаберного дыхания легочным. У наиболее примитивной из ныне живущих рептилий - гаттерии - имеются и сонные и боталловы протоки.
Центральная нервная система рептилий характеризуется возрастанием относительных размеров головного мозга и прогрессивным развитием полушарий переднего мозга, в крыше которых впервые появляется поверхностно расположенное серое вещество - кора головного мозга (развитая еще слабо).
Среди прогрессивных особенностей скелета пресмыкающихся важно появление замкнутой грудной клетки. Помимо уже указанного значения этого преобразования в интенсификации дыхания грудная клетка обеспечивает связь плечевого пояса с осевым скелетом, а тем самым - его более прочную фиксацию, что ведет к усилению опорной функции конечностей. У змей, вторично утративших парные конечности, редуцируются и их пояса.
Благодаря единственному затылочному мыщелку, специфическому устройству первых шейных позвонков, допускающему вращение атласа вокруг зубовидного отростка эпистрофея, и появлению многих шейных позвонков, подвижно сочлененных между собой, значительно увеличилась подвижность головы рептилий. Это имеет большое биологическое значение при ориентировке, отыскании и добывании пищи и т. п.
Весьма существенны эволюционные изменения в крыше черепа. У древнейших пресмыкающихся верхняя и боковые стенки черепа были составлены многочисленными покровными костями и имели вид сплошного костного панциря. Такой череп назывался стегальным. Дальнейшая эволюция шла по пути постепенной редукции покровных костей стегального черепа таким образом, что на месте редуцировавшихся костей образовывались впадины (височные ямы), ограниченные относительно тонкими костными мостиками (височными дугами). Эти преобразования черепа, с одной стороны, вели к его облегчению, что важно при обитании в значительно менее плотной, чем водная, воздушной среде; с другой стороны, углубления, образовавшиеся на месте редуцированных костей крыши черепа, были использованы. Для размещения челюстной мускулатуры. Развитие челюстной мускулатуры прямо связано с повышением активности питания.
У большинства групп пресмыкающихся в процессе редукции вторичной крыши черепа образовались две височные ямы: верхняя, ограниченная снаружи верхней височной дугой (заднелобная и чешуйчатая кости), и боковая, ограниченная сверху верхней, а снаружи нижней (скуловая и квадратно-скуловая кости) височными дугами. Рептилии с таким типом черепа относятся к группе диапсида (двудужные). У некоторых представителей этой группы редукция костей пошла дальше в направлении утраты одной или даже обеих височных дуг. Типично выраженный диапсидный череп характерен из ныне живущих форм для гаттерии и крокодилов. У многих ящериц частично редуцируется нижняя височная дуга, а у некоторых - и верхняя. У змей редуцированы обе височные дуги. От диапсидных рептилий произошли птицы (у этих животных полностью сохранилась нижняя височная дуга, но редуцирована верхняя).
У части рептилий эволюция височной области черепа пошла по пути образования только одной височной ямы, ограниченной единственной височной дугой, составленной скуловой и чешуйчатой костями. Поскольку скуловая кость - характерный элемент нижней, а чешуйчатая - верхней височных дуг диапсидного черепа, такая дуга называется смешанной, а рептилии, имеющие такое устройство черепа, относятся к группе синапсида (пресмыкающиеся со смешанной дугой). Среди современных рептилий синапсидных форм нет. Такой тип черепа характерен для потомков синапсидных пресмыкающихся - млекопитающих.
У одной из ветвей примитивных пресмыкающихся редукция покровных костей крыши черепа шла иным путем: образование выемки в затылочной части покровного черепа привело к формированию на каждой стороне одной полости - ложной височной ямы, прикрытой снаружи широким костным мостиком из заднелобной, чешуйчатой и квадратно-скуловой костей. Такие пресмыкающиеся относятся к группе анапсида (бездужные). Из ныне живущих рептилий к этой группе принадлежат черепахи.
Совокупность прогрессивных черт рептилий обусловила общее повышение энергии жизнедеятельности этих животных и появление среди них большого разнообразия жизненных форм, отражающих приспособления к различным условиям обитания. В отличие от земноводных, питающихся исключительно животной пищей, среди пресмыкающихся имеются и растительноядные формы (таковы, например, наземные черепахи). Но все же большинство современных рептилий - плотоядные животные. Большая, по сравнению с земноводными, подвижность позволяет им охотиться за подвижной добычей, иногда достигающей значительных размеров. Успеху охоты в ряде случаев способствует ряд специальных приспособлений: ядовитые зубы у некоторых видов змей, покровительственная окраска, свойственная многим рептилиям, и т. п.
Большинство рептилий - наземные животные. На суше разные виды пресмыкающихся встречаются в самых разнообразных условиях: в пустынях, зарослях болотной растительности, лесах, среди скал и т. д. Все эти виды имеют соответствующие приспособления для передвижения (окаймленные роговыми пластинками пальцы в песчаных пустынях или роговые щетинки, позволяющие лазящим формам держаться на отвесных поверхностях), чтобы избегать нападения хищников (зарывание в песок, автотомия хвоста и т. д.) и т. п.
Многие пресмыкающиеся вторично перешли к водному образу жизни (морские и пресноводные черепахи, крокодилы, водные змеи). Некоторые из этих видов почти всю жизнь проводят в воде, но обязательно выходят на сушу для размножения. Морские змеи выработали способность к яйцеживорождению, что позволило им вновь полностью перейти к водному образу жизни (среди вымерших пресмыкающихся полностью водный образ жизни вели ихтиозавры, также обладавшие способностью к яйцеживорождению).
Наибольшего разнообразия жизненных форм пресмыкающиеся достигали в период эволюционного расцвета этого класса - в мезозойскую эру. Среди ископаемых рептилий известны травоядные и хищные (разные группы динозавров), глубоко специализированные обитатели водной среды (ихтиозавры) и летающие формы (птеродактили, рамфоринхи), а также много других биологических групп. Общее похолодание климата, наступившее после мезозоя, не способствовало дальнейшему процветанию пресмыкающихся: как и амфибии, рептилии пойкилотермны и не могут долго удерживать высокую температуру тела при похолоданиях.
Большинство видов современных рептилий обитает в районах жаркого климата; виды, распространенные в умеренной зоне, значительную часть года проводят в оцепенении. В этих условиях явные преимущества оказались на стороне их «потомков» - птиц и млекопитающих, в эволюции которых появилось новое ценное качество - гомойотермность. Способность к сохранению активной жизнедеятельности в широком диапазоне температуры внешней среды - одна из главных причин, определивших господствующее положение птиц и млекопитающих в современной фауне позвоночных.
Дополнительная литература
Гуртовой Н. Н., Матвеев Б. С. Дзержинский Ф. Я. Практическая зоотомия позвоночных. Земноводные, пресмыкающиеся. М., 1978
Терентьев П. В. Герпетология. М., 1961.
Шмальгаузен И. И. Основы сравнительной анатомии позвоночных животных. М., 1947
Позвоночник объединяет 4 отдела: шейный, пояснично-грудной, крестцовый и хвостовой. Тела позвонков спереди вогнутые, сзади выпуклые — процельные позвонки.
Верхние дуги заканчиваются хорошо выраженным остистым отростком; от переднего края верхней дуги отходит пара коротких передних сочленовных отростков, а от заднего края той же дуги — пара задних сочленовных отростков. Никаких признаков остатков хорды у взрослых особей нет.
Скелет ящерицы (по Брэму):
1 — ключица, 2 — лопатка, 3 — плечо, 4 — лучевая кость, 5 — локтевая кость, 6 — запястье, 7 —пястье, 8 — фаланги пальцев, 9 — ребра, 10 — таз, 11 — бедро, 12 — большая берцовая кость, 13 — малая берцовая кость, 14 — предплюсна, 15 — плюсна, 16 — хвостовые позвонки
Шейный отдел представлен восьмью позвонками. Передний из них, носящий название атласа (atlas), или атланта, имеет характерную для всех амниот форму кольца. На передней стороне его нижней части находится непарная сочленовная поверхность, при помощи которой позвонок сочленяется с черепом. Отверстие атласа разделено связкой на две половины — верхнюю и нижнюю. Через верхнее отверстие проходит спинной мозг, а в нижнее входит зубовидный отросток (processus odontoideus), который прочно соединен, но не сращен со вторым позвонком, носящим название эпистрофея (epistropheus). Как доказывают эмбриологические данные, зубовидный отросток является отделившимся телом первого позвонка. Таким образом, атлас подвижен относительно собственного тела. Три последних шейных позвонка несут по паре хорошо развитых шейных ребер, которые подвижно причленяются к телам позвонков, постепенно увеличиваются по направлению кзади и оканчиваются свободно, не доходя до грудины. В целом шейный отдел пресмыкающихся отличается от такового земноводных не только большим количеством позвонков, но и более подвижным: сочленением первого позвонка как с черепом, так и со вторым позвонком. В результате голова получает гораздо большую свободу движений.
Пояснично-грудной отдел у ящерицы содержит 22 позвонка. Все они снабжены подвижными ребрами, длина которых постепенно уменьшается по направлению кзади. Каждое ребро состоит из верхнего костного и нижнего хрящевого отделов. Первые пять пояснично-грудных позвонков (собственно грудные позвонки) имеют особенно длинные ребра, которые причленяются к грудине, образуя грудную клетку (thorax). Грудина (sternum) хрящевая и имеет форму ромбической пластинки. Эмбриональное развитие грудины амниот показывает, что она образуется путем срастания дисталь ных концов ребер и, следовательно, не гомологична «грудине» земноводных, образующейся самостоятельным путем. Таким образом, у ящерицы уже намечается крайне характерное для амниот разделение туловищного отдела позвоночника на грудной и поясничный.
Хвостовой отдел состоит из многочисленных позвонков (несколько десятков). Передние из них похожи на крестцовые, но тела у них длиннее, поперечные отростки тоньше, а остистые—длиннее. По направлению кзади хвостовые позвонки постепенно уменьшаются в размерах, утрачивают отростки и превращаются в небольшие палочковидные косточки.
Еще интересные статьи
Развитие и усложнение двигательной системы пресмыкающихся вызвано переходом к преимущественному обитанию на суше и необходимостью преодоления неровностей её рельефа. Основные изменения опорного аппарата рептилий, по сравнению с земноводными, связаны с усилением подвижности , необходимой для успешного поиска пищи, размножения, защиты от опасности и т. д. Это нашло отражение в упрочении черепа, большей дифференцировке осевого скелета, укреплении конечностей и их поясов, дальнейшем развитии специализированной мускулатуры.
Скелет пресмыкающихся характеризуется значительным развитием костных элементов и соответствующим уменьшением хрящевых образований. Особенно большое количество покровных окостенений слагает крышу, бока и дно мозгового черепа. Существенные изменения претерпел и висцеральный отдел. Челюстной аппарат стал более массивным, кости верхней челюсти глубоко вросли в верхнюю часть мозгового отдела. Вместе с тем, череп приобрёл подвижность в соединении мозгового и висцерального отделов. Так, у змей нижняя часть черепа представляет собой сложную рычажно-шарнирную систему , позволяющую целиком заглатывать крупную добычу (удавы, яичные змеи и др.). Этому способствуют височные ямы и дуги, являющиеся местом прикрепления сильной и эластичной мускулатуры . Скелет головы сочленяется с позвоночником посредством одного мыщелка , образованного костями затылка. В целом, череп стал прочнее и, вместе с тем, легче.
Усложнение дифференцировки позвоночника проявляется в увеличении числа позвонков в отделах туловища. Особое значение имеет увеличение шейного отдела, который определяет подвижность головы, что, в свою очередь, усиливает ориентировку на местности и способствует ускорению передвижения. Путём срастания передней части рёбер с грудиной формируется грудная клетка . В области грудной клетки развивается межрёберная мускулатура , которая определяет механизм дыхания у многих видов.
Конечности большинства рептилий короткие, причленяются к поясам конечностей и имеют специализированную мускулатуру. В области соединения предплечья с кистью формируется интеркарпальное сочленение . Аналогичное образование имеет место и в задних конечностях, где в голеностопной части формируется интертарзальное сочленение . Эти образования значительно способствуют упрочению и подвижности нижних отделов конечностей. Пояса конечностей при сохранении значительного количества хрящей крепят конечности к осевому скелету .
Пресмыкающиеся, утратившие конечности, их пояса и грудную клетку (змеи, безногие ящерицы), имеют длинные рёбра, сильную туловищную и подкожную мускулатуру. У ряда рептилий (черепахи) утрата быстроты передвижения связана с развитием особого защитного приспособления – неподвижного панциря , сковывающего движения. Особенности строения скелета есть и у вторичноводных видов (морские черепахи, морские змеи, игуаны, крокодилы).
После ознакомления с основными характеристиками скелета можно перейти к более подробному рассмотрению строения его составляющих частей.
Череп
Череп вытянутой формы, со значительным содержанием костных элементов (рис. 51). Та или иная форма черепа, соотношение размеров составляющих его отделов у разных рептилий определяется типом питания и способом захвата пищи.
Мозговой череп упрочен за счёт увеличения количества костей и их прочного срастания между собой.
Крыша черепа образована парными носовыми, переднелобными, лобными, заднелобными (заглазничными) и теменными костями. Непарная межтеменная кость имеет отверстие теменного органа. К бокам этих элементов прирастают слёзные и надглазничные кости , которые обрамляют верхнюю часть орбиты глаз.
Дно черепа формируют основная клиновидная кость с рудиментарным парасфеноидом и парные сошники , по бокам от которых лежат хоаны. В образовании дна мозгового черепа принимают участие и элементы висцерального отдела. Крыловидные кости сочленяются с квадратными, которые у змей и ящериц подвижно соединены с мозговым черепом и выполняют роль подвеска для челюстного аппарата. Между крыловидными костями и верхнечелюстными находится поперечная кость , которая дополнительно укрепляет череп.
Затылочный отдел состоит из четырёх костей – основной, верхнезатылочной и парных боковых. В результате их срастания образуется один затылочный мыщелок . В нижней части отдела имеется затылочное отверстие для соединения головного и спинного мозга.
Боковую часть черепа слагают три пары ушных костей (переднеушная, верхнеушная и заднеушная), которые участвуют в образовании височных ям. Верхнеушные и заднеушные кости срастаются с костями затылочного отдела. Рядом с ними располагаются височные, чешуйчатые , верхние части крыловидных костей. Парные столбчатые кости соединяют крышу черепа с дном. Крупные глазницы формируют прилегающие костные элементы – надглазничные, заглазничные, скуловые, слёзные. Межглазничная перегородка образована выростами лобных и теменных костей. В переднюю часть мозгового черепа высоко вклиниваются верхнечелюстные и межчелюстные кости . Обонятельная область черепа хрящевая.
Рис. 51. Череп ящерицы сверху (А), снизу (Б), сбоку (В) и ее нижняя челюсть:
1 – затылочное отверстие, 2 – боковая затылочная кость, 3 – верхняя затылочная кость, 4 – основная затылочная кость, 5– затылочный мыщелок, 6 – основная клиновидная кость, 7 – сошник, 8 – хоаны, 9 – рудиментарный парасфеноид, 10 – теменная кость, 11 – отверстие для теменного глаза, 12 – лобная кость, 13 – носовая кость, 14 – межчелюстная кость, 15 – верхнечелюстная кость, 16 – предлобная кость, 17 – слезная кость, 18 – предглазничная кость, 19 – заглазничная кость, 20 – чешуйчатая кость, 21 – височная кость, 22 – скуловая кость, 23 – ноздря, 24 – квадратная кость, 25 – небная кость, 26 – крыловидная кость, 27 – столбчатая кость, 28 – поперечная кость, 29 – сочленовная кость, 30 – зубная кость, 31 – угловая кость, 32 – надугловая кость, 33 – венечный отросток
Череп ряда рептилий отличается наличием хорошо выраженных височных дуг (у ящерицы они редуцированы). У варана имеют место только верхние височные дуги , в образовании которых принимают участие заглазничные и чешуйчатые кости. Эти дуги отграничивают боковые височные ямы . Крокодилы, наряду с верхними дугами, имеют и нижние , которые формируются верхнечелюстными и скуловыми костями (диапсидный череп ) (рис. 52). У черепах отсутствуют височные ямы и височные дуги (череп анапсидного типа).
Висцеральный череп состоит изчелюстного и подъязычного аппарата . Кости верхней челюсти (межчелюстные и верхнечелюстные), как уже отмечалось выше, упрочились и глубоко вросли в мозговой череп. Имеет место аутостилия – непосредственное соединение висцерального черепа с мозговым отделом. В результате окостенения и преобразования нёбно-квадратного хряща возникли квадратные и столбчатые кости. Нёбные и крыловидные элементы – кожного происхождения и участвуют в образовании дна мозгового черепа.
У ряда рептилий (крокодилы, черепахи) имеет место вторичное костное нёбо , которое сформировалось разрастанием ряда костей – межчелюстных, верхнечелюстных, нёбных и крыловидных . У крокодилов изменилось и положение ноздрей : наружные ноздри приподняты над поверхностью головы, а хоаны отодвинуты к затылочной области черепа. Эти преобразования лежат в основе отделения носоглотки от ротовой полости, что позволяет животным держать под водой пасть открытой и одновременно дышать через ноздри, выставленные на поверхность воды.
Нижняя челюсть сформирована путём окостенения меккелева хряща и появления сочленовной, зубной, угловой, надугловой, венечной костей и ряда более мелких образований. Из перечисленных костей лишь сочленовная имеет хондральное происхождение, все остальные – покровные.
Подъязычный аппарат представляет собой пластинку (гомолог копулы ) с тремя парами рожков – производных гиоидов и жаберных дуг. Гиомандибуляре, как и у амфибий, в виде слуховой косточки (стремя) находится в среднем ухе.
Оборудование и материалы
1. Скелеты ящерицы, черепахи, змеи (по одному набору на двух студентов).
2. Позвонки ящерицы из разных отделов тела.
3. Передние и задние конечности ящерицы с поясами.
4. Череп ящерицы, черепахи, змеи.
5. Таблицы: 1) скелет ящерицы; 2) строение позвонков из разных отделов тела; 3) череп (вид сверху и снизу); 4) скелет конечностей и их поясов.
Вводные замечания
Основным направлением эволюции пресмыкающихся было совершенствование локомоторного аппарата и увеличение разнообразия движений. Пресмыкающиеся (современные и вымершие) демонстрируют все принципиальные способы передвижения, известные для позвоночных.
Эволюционно исходный и типичный способ передвижения сохранился у современных ящериц. Как и у хвостатых земноводных, перемещение осуществляется путем поочередной перестановки относительно коротких расставленных в стороны конечностей
при изгибании туловища вправо - влево. Строение суставов конечностей и общая схема локомоции у современных ящериц лишь слегка видоизменена по сравнению с ранними пресмыкающимися.
Эффективность такого способа передвижения у пресмыкающихся стала выше по сравнению с земноводными. Это было достигнуто, с одной стороны, усилением и дифференциацией мускулатуры тела и конечностей, а с другой стороны, дифференциацией скелета и укреплением и преобразованием отдельных его узлов и сочленений. Усиление мышц конечностей, развитие продольной мускулатуры тела, смещение в той или иной мере конечностей под туловище - все это сделало возможным быстрое перемещение с подъемом тела над землей (бег). Усиление мускулатуры и перемещение в разнообразной среде сопровождалось увеличением нагрузки на скелет. Это потребовало его укрепления. Поэтому грубоволокнистая ткань трубчатых костей сменилась на тонковолокнистую, ключицы стали играть роль своеобразной распорки между дорсальной (дистальный край лопатки) и вентральной (грудина) частями плечевого пояса. Наиболее распространенным стал процельный тип позвонков. Все туловищные позвонки, а также часть шейных и хвостовых несут подвижные ребра. Интенсивные движения сгибания - разгибания кисти и стопы стали возможными благодаря появлению интеркарпального и интертарзального суставов.
Пресмыкающимся свойственна большая подвижность головы. Это обеспечено чрезвычайно подвижным сочленением черепа с шейным отделом позвоночника: во-первых, череп соединяется с первым шейным позвонком одним мыщелком; во-вторых, первые два шейных позвонка (атлант и эпистрофей) видоизменены так, что возможны движения головы вокруг продольной оси тела. Подвижность головы и шеи в сочетании со способностью к быстрому перемещению по субстрату обеспечили пресмыкающимся широкие возможности для поиска пищи. Сильное сжатия челюстей оказалось важным для захвата разнообразной добычи. В связи с этим развилась мощная жевательная мускулатура и произошла перестройка крыши и боков черепа - появились височные ямы, что значительно увеличило поверхность для прикрепления челюстных мышц. Деформация черепа при сжатии жевательных мышц предотвращается столбчатыми костями.
Соединение ребер с грудиной и формирование таким образом грудной клетки - еще одно крупное преобразование скелета пресмыкающихся. В результате резко увеличилась скорость вентиляции легких и возрос объем прокачиваемого воздуха. Это способствовало отказу от кожного дыхания и повышению интенсивности газообмена.
В целом скелет пресмыкающихся несет комплекс черт, которые демонстрируют более высокий уровень организации этих животных и более глубокие адаптации к жизни на суше.
Изучите скелет пресмыкающихся.
Рассмотрите :
Череп
Мозговой череп
Затылочный отдел: верхняя; основная и две боковые затылочные кости; затылочное отверстие; затылочный мыщелок.
Бока черепа: скуловая; чешуйчатая; крыловидная; столбчатая; квадратная; ушные кости.
Крыша черепа: носовые; предлобные; лобные; теменные; межтеменная; слезные; надглазничные; заглазничные кости.
Дно черепа: основная клиновидная кость; сошник; зачатки вторичного костного нёба.
Висцеральный череп
Верхняя челюсть: верхнечелюстные; межчелюстные кости.
Нижняя челюсть: зубные; сочленовные; угловые; надугловые кости;
Подъязычный аппарат: подъязычная кость с двумя парами рожков.
Позвоночник
Отделы позвоночника: шейный; грудной; поясничный; крестцовый; хвостовой; туловищный позвонок; атлант; эпистрофей. Грудная клетка: грудина; ребра.
Пояса конечностей
Плечевой пояс: грудина; надгрудинник; коракоиды; лопатки; ключицы.
Тазовый пояс: подвздошные; седалищные; лобковые кости; вертлужная впадина.
Парные конечности
Передняя конечность: плечо; предплечье (локтевая и лучевая кости); запястье; пясть; фаланги пальцев; интеркарпальный сустав.
Задняя конечность: бедро; голень (большая и малая берцовые кости); предплюсна; плюсна; фаланги пальцев; интертарзальный сустав.
Зарисуйте :
1) череп (вид сверху, сбоку, снизу); 2) пояс передних конечностей; 3) тазовый пояс и крестцовый отдел позвоночника.
Строение скелета
Мозговой череп
У ящериц череп почти полностью окостеневший и состоит из большого количества костей, образующих довольно массивную мозговую коробку (рис. 70, 71).
Затылочный отдел черепа состоит из четырех затылочных костей, ограничивающих затылочное отверстие. Основная затылочная кость совместно с двумя боковыми затылочными костями образует затылочный мыщелок, который входит в сочленовную ямку атланта. Верхняя затылочная кость в виде арки прикрывает сверху затылочное отверстие и заходит сзади под крышу черепа.
Бока черепа в области слуховой капсулы сформированы тремя ушными костями, которые участвуют в образовании височных ям.
Рис. 70. Строение черепа ящерицы:
А
- вид сверху; Б
- вид снизу; В
- вид сбоку; 1
- затылочное отверстие; 2
- боковая затылочная кость; 3
- верхняя затылочная кость; 4
- основная затылочная кость; 5
- затылочный мыщелок; 6
- основная клиновидная кость; 7
- сошник; 8
- хоаны; 9
- парасфеноид; 10
- теменная кость; 11
- отверстие для теменного органа; 12
- лобная кость; 13
- носовая кость; 14
- межчелюстная кость; 15
- верхнечелюстная кость; 16
- предлобная кость; 17
- слезная кость; 18
- надглазничные кости; 19
- заглазничная (заднелобная) кость; 20
- чешуйчатая кость; 21
- височная кость; 22
- скуловая кость; 23
- ноздря; 24
- квадратная кость; 25
- нёбная кость; 26
- крыловидная кость; 27
- столбчатая кость; 28
- поперечная кость; 29
- сочленовная кость; 30
- зубная кость; 31
- угловая кость; 32
- надугловая кость; 33
- венечная кость

Рис. 71. Строение черепа варана:
А
- вид сверху, Б
- вид снизу, 1
- межчелюстная кость, 2
- носовая кость, 3
- предлобная кость, 4
- лобная кость, 5
- слезная кость, 6
- надглазничная кость, 7
- теменная кость, 8
- верхняя затылочная кость, 9
- основная затылочная кость, 10
- боковая затылочная кость, 11
- чешуйчатая кость, 12
- квадратная кость, 13
- заглазничная (заднелобная) кость, 14
- скуловая кость, 75 - верхнечелюстная кость, 16
- сошник, 17
- небная кость, 18
- поперечная кость, 19
- крыловидная кость, 20
- основная клиновидная кость, 21
- затылочное отверстие, 22
- квадратно-скуловая кость, 23
- затылочный мыщелок
Переднеушные кости хорошо развиты и обособлены, верхнеушные кости срастаются с верхней затылочной костью, а заднеушные - с боковыми затылочными костями. На боках черепа располагаются большие глазницы. Межглазничная перегородка образована отростками лобных и теменных костей крыши черепа. Позади глазницы лежат заглазничные, скуловые и чешуйчатые кости. В переднем углу глазницы расположены небольшие слезные кости, а ряд мелких надглазничных косточек окаймляет глазницу сверху. Переднюю часть черепа формируют производные висцерального черепа - верхнечелюстные и межчелюстные кости. В обонятельной области у ящерицы окостенений нет, обонятельная капсула с наружными ноздрями образована хрящем.
Крышу черепа формируют парные носовые, предлобные, лобные и теменные кости Между теменными костями лежит непарная межтеменная кость с отверстием для теменного органа.
Дно черепа образовано основной клиновидной костью, с которой срастается рудиментарный парасфеноид. Спереди от парасфеноида лежат небные кости и сошники, по бокам от которых
находятся хоаны. В формировании дна мозговой коробки участвуют также некоторые кости висцерального черепа - это нёбные и крыловидные кости. Изогнутые крыловидные кости передними концами упираются в нёбные кости, а задними концами сочленяются с основной клиновидной и с квадратными костями.
В височной области у ящериц расположена височная яма, ограниченная единственной верхней височной дугой, которая образована заглазничной и чешуйчатой костями. Нижняя височная дуга у ящериц редуцирована. В глубине височных ям хорошо заметны столбчатые (надкрыловидные) кости в виде небольших стоек, упирающихся в крыловидные и теменные кости.
Висцеральный череп
Челюстная дуга у ящериц сильно видоизменена. В результате окостенения нёбно-квадратного хряща у них сформировались квадратные, крыловидные, столбчатые (надкрыловидные) и нёбные кости. Квадратная кость верхним концом подвижно сочленяется с чешуйчатой костью, а нижним - с нижней челюстью и служит для нее подвеском. Появление сустава между чешуйчатой костью и верхним концом квадратной кости позволило увеличить силу сжатия челюстей и, по-видимому, стало причиной эволюционного успеха ящериц. Верхняя челюсть образована верхнечелюстными и межчелюстными костями, на которых располагаются зубы. Между крыловидными и верхнечелюстными костями лежат поперечные кости, которые выступают в качестве распорок и укрепляют череп.
Основой для развития костей нижней челюсти служит меккелев хрящ. Он замещается сочленовной костью, сочленяющейся с квадратной костью. Кроме того, в состав нижней челюсти входит ряд покровных костей: самые крупные зубные кости, которые несут зубы, и более мелкие - венечная, угловая и надугловая.
Верхний отдел подъязычной дуги - гиомандибуляре - входит в состав среднего уха в виде слуховой косточки - стремечка. Остальная часть подъязычной дуги и остатки жаберных дуг преобразуются в подъязычный аппарат, представленный хрящевой подъязычной пластинкой и тремя парами рожек.
Позвоночник
Позвоночник пресмыкающихся подразделяется на четыре отдела: шейный, грудной, поясничный, крестцовый и хвостовой .
У большинства пресмыкающихся позвонки процелъные , т.е. передневогнутые. Тело позвонка удлиненное, спереди вогнутое,

Рис. 72. Два первых шейных позвонка варана:
А
- атлант; Б
- эпистрофей; 1
- верхняя дуга; 2
- спинно-мозговой канал; 3
4
- зубовидный отросток эпистрофея; 5
- тело позвонка
верхняя дута заканчивается верхним остистым отростком. От тела позвонка отходят передние и задние сочленовные отростки. У основания верхней дуги находятся сочленовные поверхности для прикрепления ребер.
Первые два шейных позвонка сильно отличаются от остальных. Первый шейный позвонок - атлант - имеет вид кольца, разделенного плотной связкой на верхнюю и нижнюю половины (рис. 72). На передней поверхности атланта имеется сочленовная ямка для затылочного мыщелка. Через верхнее отверстие головной мозг соединяется со спинным, а в нижнее отверстие сзади входит зубовидный отросток второго шейного позвонка - эпистрофея . Такое сочленение черепа с шеей обеспечивает сложные движения головы. Задние шейные позвонки несут ребра.
К позвонкам грудного отдела причленяются ребра , которые сочленяются с грудиной и формируют грудную клетку (истинные ребра). Поясничные позвонки тоже несут ребра, которые не соединяются с грудиной (ложные ребра). Два крестцовых позвонка имеют массивные поперечные отростки, которые сочленяются с тазовым поясом (см. рис. 75). Количество хвостовых позвонков у ящериц варьирует, достигая нескольких десятков. Передние хвостовые позвонки несут поперечные и остистые отростки (рис. 73), а к концу хвоста позвонки преобразуются в палочковидные косточки. Тела почти всех хвостовых позвонков разделены тонкой хрящевой прослойкой, по которой происходит разлом при автотомии.

Рис. 73. Строение переднего хвостового позвонка ящерицы:
А
- вид сверху; Б
- вид сбоку; 1
- тело позвонка; 2
- верхний остистый отросток; 3
- передний сочленовный отросток; 4
- задний сочленовный отросток; 5
- поперечный отросток; 6
- нижний остистый отросток; 7
- нижняя дуга
Пояса конечностей
Пояс передних конечностей у ящериц (рис. 74) устроен в целом сходно с плечевым поясом земноводных, однако по сравнению с ним здесь сильно развиты окостенения. В месте соединения коракоида с лопаткой имеется суставная впадина для сочленения с плечевой костью. К дорсальному краю лопатки прирастает надлопаточный хрящ, а спереди к коракоиду - прокоракоидный хрящ. Коракоид и прокоракоид срастаются, образуя единый скелетный элемент, который сочленен с грудиной подвижно. В этом суставе возможны скользящие движения в горизонтальной плоскости вдоль переднего края грудины, это увеличивает длину шага. Снизу к грудине прирастает крестообразный надгрудинник покровного происхождения. Ключицы соединяют дорсальный край лопатки с надгрудинником. Таким образом, в отличие от земноводных ключицы пресмыкающихся ориентированы в вертикальной плоскости и делают плечевой пояс более прочным.
Тазовый пояс у ящериц хорошо развит и состоит из двух безымянных костей, которые образовались путем слияния тазовых костей - подвздошной, седалищной и лобковой (рис. 75). Имеются суставные впадины для головки бедренной кости (вертлужные впадины). Правые и левые лобковые и седалищные кости соединяются хрящевыми симфизами. Тазовый пояс прочно соединен с позвоночником путем прирастания подвздошных костей к поперечным отросткам крестцовых позвонков.
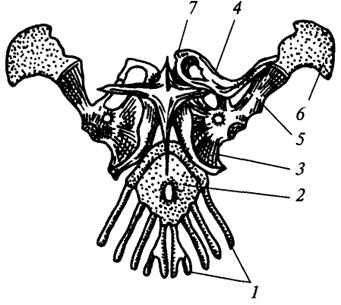
Рис. 74. Плечевой пояс ящерицы:
1
- первые четыре грудных ребра; 2
- грудина; 3
- коракоид; 4
- ключица; 5
- лопатка; 6
- надлопаточный хрящ; 7
- надгрудинник

Рис. 75. Тазовый пояс ящерицы:br1
- суставная (вертлужная) впадина для головки бедра; 2
- подвздошная кость; 3
- лобковая кость; 4
- седалищная кость; 5
- симфиз
Парные конечности
Конечности пресмыкающихся соответствуют общей схеме строения пятипалой конечности наземных позвоночных. Скелет передней конечности включает плечевую кость, локтевую и лучевую кости, два ряда мелких костей запястья, ряд костей пясти и ряд костей фаланг пальцев. Скелет задней конечности включает бедренную кость, большую и малую берцовые кости, два ряда мелких костей предплюсны, ряд костей плюсны и ряд костей фаланг пальцев. К особенностям строения конечностей следует отнести формирование интеркарпального (в передней конечности) и ин- тертарзального (в задней конечности) суставов (рис. 76).

Рис. 76. Строение интертарзального сустава ящерицы (правая стопа прыткой ящерицы сверху):
1
- сросшиеся проксимальные косточки предплюсны; 2
- дистальные косточки предплюсны; 3
- большая берцовая кость; 4
- малая берцовая кость; 5
- кости плюсны
Интеркарпальный сустав формируется между двумя рядами косточек запястья, интертарзальный сустав - между двумя рядами косточек предплюсны. Эти суставы обеспечивает возможность сильного сгибания и разгибания кисти и стопы, что важно при своеобразном типе локомоции.