Câte vertebre sacre are o șopârlă. Sistemul musculo-scheletic al reptilelor caracteristicile generale ale scheletului
TEMA 14. SKELETONUL UNUI REBUMAN
POZIȚIA SISTEMATICĂ A OBIECTULUI
Subtipul Vertebratelor, Vertebratelor
Reptilia Class, Reptilia
1. Scara de echipă, Squamata
Reprezentant - Varan Gray, Varanus griseus Daud.
2. Crocodilii din echipa, Crocodilia
Reprezentant - Mississippi Alligator, Alligator mississippiensts Daud.
3. Turtle Squad, Chelonia (Testudines)
Reprezentant - țestoasa de mlaștină, orbicular Etnys este L.
MATERIAL ȘI ECHIPAMENT
Scheletele montate de șopârlele de monitor, șerpi, țestoase - unul pe grup de studenți; cranii de crocodil (sau manechine) - 1-2 per grup. Pentru unul sau doi studenți sunt necesari:
1. Scheletul pliabil al unei șopârle monitor.
2. Craniul țestoasei.
3. Ace de disecție - 2.
JOB
Luați în considerare scheletul unui șopârlă monitor în ansamblu. Găsiți-i principalele părți: craniul, coloana vertebrală, scheletul membrelor împerecheate și centurile lor. Luați în considerare caracteristicile structurale ale scheletului unui șarpe și a unei țestoase. Luați în considerare detaliile structurii scheletului unui șopârlă monitor și a craniilor unui șopârlă monitor, crocodil și țestoasă. Comparați structura craniului unui crocodil, monitorizați șopârlă, șarpe și broască țestoasă.
Efectuați următoarele desene:
1. Craniul șopârlei monitor din lateral.
2. Craniul șopârlei monitor de jos.
4. Prelungirea unei șopârle monitor.
6. Membrul posterior al șopârlei monitor.
7. Craniu de crocodil deasupra.
8. Craniul de crocodil din fund.
9. Craniul țestoasei din lateral.
DESCRIEREA SKELETONULUI
Schelet axial. Diferențierea scheletului axial, sau a coloanei vertebrale, în secții este mai accentuată în reptile decât în \u200b\u200bamfibieni. Coloana vertebrală cervicală (pars cervicalis) este întotdeauna compusă din mai multe vertebre, dintre care cele două anterioare au o structură specială. Prima vertebră cervicală se numește atlas sau atlas (atlas; Fig. 81, A). Este lipsit de corpul vertebral și are forma unui inel împărțit în două părți. Pe suprafața frontală inferioară a acestei vertebre există o cavitate articulară, conectată mobil cu condilul craniului (vezi mai jos). A doua vertebră cervicală - epistrofie (epistrofă; Fig. 81, B), are în față un proces asemănător unui dinte (procese odontoideus, fig. 81, 1), care este corpul primei vertebre cervicale fuzionată cu epistrofie. Procesul asemănător unui dinte intră liber în deschiderea inferioară a atlasului. Această structură a primelor vertebre cervicale asigură o mobilitate mai mare a capului. Vertebrele cervicale rămase au dispozitivul obișnuit (vezi mai jos); multe dintre ele au coaste scurte ale gâtului.
Fig. 81. vertebrele vertebrale
A - atlas; B - o epistrofie; În - o vertebră toracică; G - secțiunea longitudinală a vertebrei toracice:
1 - proces de epistrofie asemănător unui dinte, 2 - corp vertebral, 3 - arc superior, 4 - proces spinos, 5 - canal pentru măduva spinării, 6 - proces articular anterior, 7 - proces articular posterior
Diviziunile toracice și lombare nu sunt destul de distincte și sunt de obicei considerate ca o singură diviziune (pars toracolumbalis). De fapt, regiunea toracică este partea coloanei vertebrale în care coastele care se extind de la vertebre cu capătul inferior se atașează de stern. Vertebrele regiunii lombare poartă coastele care nu ajung la stern. Corpurile vertebrale (Fig. 81, 2) sunt concave în față și în spate convexe; astfel de vertebre se numesc procent. Deasupra corpului vertebral, arcadele superioare se ridică (arcus neuralis; Fig. 81, 3), încheindu-se cu un proces spinos (processusspinosus; Fig. 81, 4). În canalul format din arcurile superioare este situat măduva spinării.
Procesele anterioare (procese articulare anterioare; Fig. 81, 6) și posterioare (procese articulare posterioare; Fig. 81, 7) procesele articulare se depărtează de părțile anterioare și posterioare ale bazei arcurilor superioare. Aceste procese asociate se conectează la procesele articulare ale vertebrelor vecine și contribuie la o mai mare rezistență a coloanei vertebrale în timpul îndoirii. Pe părțile laterale ale corpului vertebral (în apropierea bazei arcadelor superioare) există mici indentări la care se atașează coastele.
Secțiunea sacrală (pars sacralis) este formată din două vertebre, care se caracterizează prin procese transversale puternic dezvoltate (proces transversus); ele sunt unite de oasele pelvisului. Secțiunea cozii (pars caudalis) este reprezentată de numeroase vertebre, scăzând treptat ca mărime.
Această structură a coloanei vertebrale este tipică clasei de reptile, dar în unele grupuri suferă modificări secundare. În special, la șerpi, în legătură cu reducerea membrelor împerecheate și apariția unui alt tip de mișcare - târâre pe abdomen prin îndoirea trunchiului - coloana vertebrală este divizată în mod clar numai în trunchiul și secțiunile cozii. Toate vertebrele trunchiului au coaste mobile, ale căror capete inferioare sunt libere (sternul șerpilor este absent) și se prind împotriva scuturilor cornei abdominale.

Fig. 82. Scheletul unei broaste țestoase
A - carapace; B - plastron:
1 - secțiunea trunchiului coloanei vertebrale, 2 - plăci costale, 3 - plăci marginale, 4 - coracoid, 5 - scapula, 6 - ilium7 - os pubian; 8 - os ischial
La țestoase, scheletul axial ia parte la formarea bazei osoase a carapaceului lor. Scutul superior al cochiliei - carapace (caparax; Fig. 82, A) - este compus din mai multe rânduri de plăci osoase. Rândul mijlociu (nepereche) al acestor plăci este format din fuziunea proceselor spinoase și transversale extinse și aplatizate ale vertebrelor trunchiului cu oasele pielii; pe părțile laterale ale rândului de mijloc sunt asezate rânduri de plăci osoase topite cu nervuri lărgite. Marginea carapaceului este formată din plăci osoase de origine integumentară. Astfel, coloana vertebrală a torsului este fixată și solidă fuzionată cu scutul dorsal al carapaceului. Coloana vertebrală cervicală și caudală sunt mobile. În același timp, vertebrele cervicale frontale sunt opistocelulare (corpul vertebral este convex în față și concavă în spate), cele posterioare sunt longitudinale, iar între aceste două grupuri există o vertebră al cărei corp are o suprafață convexă atât în \u200b\u200bfață cât și în spate.
Craniu. Comparativ cu amfibienii, craniul reptilelor se caracterizează printr-o osificare mult mai completă. O anumită cantitate de cartilaj se păstrează numai în capsula olfactivă și în regiunea auditivă. Părțile axiale și viscerale ale craniului sunt embrionate așezate separat, dar la animalele adulte reprezintă o singură entitate. Compoziția craniului include atât cartilaj (înlocuitor sau primar), cât și numeroase oase ale pielii (integumentare sau secundare). Este convenabil să folosiți craniul unei șopârle mari, șopârlă monitor, ca obiect principal pentru studiu.

Fig. 83. Craniul șopârlei monitor
A este pe partea laterală; B - de jos; B - de sus; G - înapoi:
1 - os occipital principal, 2 - os occipital lateral, 3 - os occipital superior, 4 - foramen occipital mare, 5 - condil occipital, 6 - os anterior, 7 - principal os sfenoid, 8 - deschizător, 9 - os parietal, 10 - os frontal, 11 - os nazal, 12 - os prefrontal, 13 - os preorbital, 14 - os lacrimal, 15 - fosa temporală superioară, 16 - os posterior, 17 - os scalos , 18 - osul maxilar, 19 - osul maxilar, 20 - osul zigomatic, 21 - ruperea arcului temporal inferior datorită reducerii osului pătrat-zigomatic, 22 - os pătrat, 23 - os pterigoid, 24 - os palatin, 25 - os supra-pterigoid, 26 - os transvers, 27 - os arcuit, 28 - os dinte, 29 - os unghiular, 30 - os articular, 31 - os coronarian
Craniu axial. În regiunea occipitală a craniului, există toate cele patru oase occipitale: principalul occipital (basioccipitale, fig. 83, 1), două occipitale laterale (occipitale laterale; fig. 83, 2) și occipitalul superior (supraoccipitale; fig. 83, 3). Aceste oase de origine primară înconjoară mare foramen occipital (foramen occipitale magnum; Fig. 83, 4). Oasele occipitale inferioare și laterale formează împreună unicul condil occipital (spre deosebire de amfibieni) (condylus occipitalis; Fig. 83, 5), care se articulează cu prima vertebră cervicală - atlasul. Articulația capului cu gâtul folosind un singur condil, în combinație cu caracteristicile structurale ale primelor două vertebre cervicale deja luate în considerare, conferă capului reptilei o mobilitate semnificativă.
În regiunea auditivă a oaselor cartilajului, doar osul anterior pereche (prooticum; Fig. 83, 6) rămâne independent, în timp ce fuziunea urechii superioare cu osul occipital superior, iar urechea posterioară cu occipitalul lateral.
Septul interorbital al reptilelor este subțire, membranos și numai crocodilii și șopârlele au mici osificări separate, aparent corespunzând oaselor în formă de pană. Capsula olfactivă nu are osificare.
La baza craniului, în fața osului occipital principal, se află un os sfenoid principal integumentar destul de mare (basefhenoideum; Fig. 83, 7). Procesul său îngust anterior este omolog cu parafenoidul (parafenoideul), care este vizibil redus în reptile. În partea anterioară a fundului craniului, sub secțiunea olfactivă, există un deschizător împerecheat (voma; Fig. 83, 8), de asemenea, de origine integumentară.
Acoperișul craniului este reprezentat de numeroase oase integumentare, unele căzând în jos și acoperă craniul din părțile laterale. Acestea includ oasele parietale (parietale; Fig. 83, 9), frontale (frontale; Fig. 83, 10) și oasele nazale (nazale; Fig. 83, 11). În fața oaselor frontale, de obicei, sunt localizate oasele prefrontal (praefrontale; Fig. 83, 12) și preorbitale (praeorbitale; Fig. 83, 13), iar sub ele în peretele anterior al orbitei se împerechează lacrimal (lacrimale; Fig. 83, 14) ).
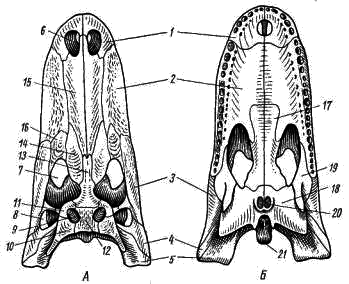
Fig. 84. Craniu de crocodil (aligator de Mississippi).
A - de sus; B - partea de jos:
1 - os maxilar, 2 - os maxilar, 3 - os zigomatic, 4 - os pătrat zigomatic, 5 - os pătrat, 6 - nară externă, 7 - orbită, 8 fosa temporală laterală, 9 - fosa temporală superioară, 10 - os scalos, 11 - osul frontal (postorbital), 12 - osul parietal, 13 - osul frontal, 14 - osul prefrontal, 15 - osul nazal, 16 - osul lacrimal, 17 - osul palatin, 18 - osul pterigoid, 19 - osul transvers , 20 - choans (deschideri interne ale nărilor), 21 - condil occipital
Dintre oasele rămase integumentare ale craniului axial, oasele care participă la formarea așa-numitelor arcuri temporale prezintă un interes deosebit. Crocodilul din acoperișul craniului din afara osului parietal (Fig. 84, 12) are o gaură pe fiecare parte - fosa temporală superioară (Fig. 84, 9). La marginea exterioară, fosa temporală superioară este delimitată de oasele frontale sau postorbitale posterioare (postfrontale-postorbitale, Fig. 84, 11) și de scalsoase (squamosum; Fig. 84, 10). Aceste două oase alcătuiesc un arc temporal superior. Pe partea craniului din spatele orbitei există gropi temporale laterale (Fig. 84, 8), delimitate extern de arcadele temporale inferioare. Fiecare arc temporal inferior este alcătuit din două oase: zigomatică (jugală; Fig. 84, 3) și pătrat-zigomatic (quadrato-jugală; Fig. 84, 4). Arcul temporal inferior se conectează la maxilarul superior: osul zigomatic crește până la maxilar, iar osul pătrat-zigomatic crește pătrat. Acest tip de craniu, ca un crocodil - cu două gropi temporale și două arcade temporale, se numește diapsid (cu două brațe).
Într-o șopârlă monitoră, fosa temporală superioară (Fig. 83, 15) este delimitată de arcul temporal complet superior (postfrontale - squamosum; Fig. 83, 16-17). Ca parte a arcului temporal inferior, osul pătrat-zigomatic a fost redus și s-a păstrat doar pomețul (Fig. 83 , 20); prin urmare, fosele temporale laterale nu sunt deschise din exterior și rămân deschise. Prin urmare, craniul șopârlei monitor poate fi considerat ca un craniu de tip diapsid, dar cu un arc inferior redus. În unele alte șopârlele, arcul temporal superior este parțial redus, în timp ce la șerpi (vezi Fig. 86) ambele arcade temporale sunt reduse (oasele frontale și scaltoase posterioare nu se conectează între ele, ambele gropi temporale rămân deschise în exterior).
Astfel, șerpii și șopârlele (squamous squad, Squamata) din structura craniului aparțin grupului de reptile diapside (cu două brațe), dar se caracterizează printr-un grad diferit de reducere a arcadelor temporale.

Fig. 85. Craniul unei broaște țestoase:
1 - fosă temporală falsă, 2 - os maxilar, 3 - os maxilar, 4 - os zigomatic, 5 - os zigomatic pătrat, 6 - os pătrat, 7 - os scalos, 8 - os frontal, 9 - os parietal, 10 - os frontal, 11 - os prefrontal, 12 - os occipital superior
Testoasa are ambele gropi temporale absente, iar peretele lateral al acoperișului craniului, delimitând cavitatea mare din exterior - așa-numita groapă temporală falsă (Fig. 85, 1), care s-a format ca o adâncitură în partea occipitală a craniului, este compusă din oase dens topite: posterior (postfrontale; Fig. 85, 8), solzoasă (squamosum; fig. 85, 7), zigomatică (jugală; fig. 85, 4) și pătrat-zigomatică (quadrato-jugale, fig. 85, 5). Acest tip de craniu, lipsit de adevăratele fose temporale și arcadele temporale care le limitează, se numește anapsidic (neînsuflețit).
Craniu visceral. În șopârlă de monitor, cartilajul palatin-pătrat se osifică, formând un os pătrat în partea posterioară (cvadrat; Fig. 83, 22), la capătul inferior de care este atașat maxilarul inferior; capătul superior al osului pătrat este articulat mobil cu un craniu axial. În fața osului pătrat se află pterigoidul (pterygoideum; fig. 83, 23), iar în fața acestuia se află osul palatin (palatin; fig. 83, 24), care se conectează la oasele maxilare și voma. Toate aceste oase sunt împerecheate; dintre acestea, doar oasele pătrate au origine cartilaginoasă (primară).
Din osul pterygoid, pterygoidul superior (epipterygoideum; Fig. 83, 25) se extinde în sus. Acest os împerecheat care leagă oasele pterygoid și parietal este omolog cu procesul vertical („ascendent”) al cartilajului pătrat palatin și este caracteristic reptilelor vii pentru șopârlele și gatteria. În plus față de pterygoidul superior, oasele transversale (transversum; Fig. 83, 26) se extind din oasele pterygoid, care în partea lor din față se alătură oaselor maxilare. Maxila secundară este reprezentată de oasele maxilare (praemaxillare; Fig. 83, 18) și maxilare (maxilare; Fig. 83, 19). Maxilarul inferior este format din osul articular primar (articulare; Fig. 83, 30) și oase integumentare: dinte (dentale; Fig. 83, 28), unghiular (angulare; Fig. 83, 29), supra-unghiular (supraangulare; Fig. 83, 27), coronare (coronare; fig. 83, 31) și, uneori, mai multe oase mai mici.
Pe oasele premaxilare, maxilar și dinte ale reptilelor (cu excepția țestoaselor) există dinți simpli conici, uneori ușor îndoit, care cresc până la marginea osului corespunzător.
Arcul sublingual, ca cel al amfibienilor, a pierdut complet funcția suspensiei. Elementul superior al arcului hioid (hyomandibular) face parte din urechea medie, sub forma unui osic auditiv în formă de tijă - scăpările (stapes seu columella), iar restul acestuia, împreună cu resturile arcadelor branhiale anterioare, formează aparatul hioid.
Structura descrisă a craniului visceral este în general tipică tuturor reptilelor. Dar în unele grupuri există abateri de la această schemă, în principal legate de specificul biologiei acestor grupuri.

Fig. 86 Craniul unui șarpe otrăvitor:
1 - osul maxilar, 2 - osul maxilar. 3 - os palatin, 4 - os pterygoid, 5 - os transvers, 6 - os pătrat, 7 - os scalp, 8 - os posterior, 9 - dinte otrăvitor, 10 - os frontal, 11 - os nazal, 12 - os dentar. 13 - os unghiular, 14 - os articular
La șerpi, nu numai pătrate (Fig. 86, 6), dar și solzoase (Fig. 86, 7) conectate cu ele, precum și oasele pterygoid (Fig. 86, 4) și oasele palatine (Fig. 86, 3) sunt foarte mobile. Ultimele două au dinții ascuțiți. Oasele transversale (Fig. 86, 5) la șerpi servesc ca pârghii, așa cum a fost, transmitând mișcarea oaselor pterigoide către oasele maxilare (Fig. 86, 2), care la rândul lor sunt foarte mobile. Acest întreg sistem de oase articulate mobil contribuie nu numai la o deschidere extrem de largă a gurii, dar oferă și mișcări independente ale jumătăților drepte și stângi ale aparatului maxilar atunci când împinge prada în gât cu interceptare alternativă. Aceasta permite șerpilor să înghită o pradă relativ mare (care depășește grosimea corpului șarpelui). Șerpii veninoși de pe oasele maxilare au atașați dinți otrăvitori, curbați spre spate, curbați înapoi (Fig. 86, 9) care au un canal intern sau o canelură pe suprafața anterioară, prin care curge veninul din glandele otrăvitoare situate la baza dintelui atunci când este mușcat.
Craniul de crocodil (Fig. 84) se caracterizează prin faptul că dinții nu cresc până la marginea oaselor dentare, maxilare și maxilare, ca în alte reptile, ci se așează în cavități speciale (găuri sau alveole) ale acestor oase - dinți tecodont. O altă caracteristică a craniului visceral al crocodililor este un palat dur secundar care separă cavitatea bucală de pasajul nazofaringian.
Procesele palatine ale oaselor maxilare (Fig. 84.7) și ale maxilarului (Fig. 84, 2), precum și ale oaselor palatine (Fig. 84, 17) și pterygoid (Fig. 84, 18) participă la formarea palatului dur secundar. Datorită formării unui palat dur, coanele secundare (Fig. 84, 20) sunt transportate înapoi și localizate în oasele pterygoid, deasupra laringelui. Formarea unui palat dur secundar este legată de natura stilului de viață al crocodilului: contactul direct al laringelui cu coanul deschide posibilitatea respirației neîntrerupte la mâncare și când crocodilul se sprijină în apă superficială, expunând nările ridicate de la apă, în timp ce cavitatea bucală este umplută cu apă.
Membrele pereche și centurile lor. Brâu de umăr reptilele constau din oase tipice: scapula localizată mai mult dorsal (scapula; Fig. 87, 1) și coracoidul (coracoideum; Fig. 87, 3) orientate spre partea ventrală. Ambele oase sunt implicate în formarea fosei articulare (Fig. 87, 4) pentru a atașa antecedentul. Dorsala spre scapula este un cartilaj suprascapular aplatizat larg (cartilago suprascapularis; Fig. 87, 2), iar în fața cortexului este procoracoid cartilaginos (cartilago procoracoidea; Fig. 87, 5). Există un stern bine dezvoltat (stern, Fig. 87, 6), la care sunt atașate mai multe coaste (costa, Fig. 87, 7). Astfel, spre deosebire de amfibieni, reptila dezvoltă un piept, iar brâul de umăr are sprijin în scheletul axial. Pe partea ventrală a sternului se află un os integumentar în formă de T - suprasternum (epistern; Fig. 87, 8), în fața acestuia - și oase integumentare - claviculă (clavicula; Fig. 87, 9). Capetele exterioare ale claviculei sunt atașate de omoplați, iar interiorul contopit cu ramurile toracului. Claviculele și suprasternul (absente la amfibieni) cresc rezistența legăturii părților drepte și stângi ale brâului de umăr.

Fig. 87. Brâul de umăr al șopârlei monitor (vedere de jos):
1 - scapula, 2 - cartilaj suprascapular, 3 - coracoid, 4 - cavitatea articulară a capului umărului, 5 - cartilaj procoracoid, 6 - stern, 7 - coaste, 8 - suprasternum, 9 - claviculă
La șerpi, brâul de umăr este complet redus, iar în țestoase, claviculele și suprasternul sunt incluse în oasele scutului abdominal (plastron; vezi Fig. 82, B) al carapaceului, formând respectiv plăcile osoase anterioare pereche și nepereche.

Fig. 88. Brâu pelvin al șopârlei monitor (vedere de jos):
1 - ileum, 2 - os pubian, 3 - os sciatic, 4 - acetabulum (fosa articulară) pentru capul femural, 5 - vertebre sacrale
Brâul pelvin este format din două jumătăți simetrice conectate de-a lungul liniei medii a cartilajului. Fiecare jumătate este formată din trei oase; localizat dorsal iliac (Ilium; Fig. 88, 1), situat pe partea ventrală a pubianului (pubis; Fig. 88, 2) și sciatic (ischium, Fig. 88, 3). Toate aceste oase participă la formarea fosei articulare (acetabulum; Fig. 88, 4), de care este atașat membrul posterior. Pelvisul reptilei este închis: oasele pubiene și ischiale dreapta și stânga de pe partea ventrală sunt contopite.

Fig. 89. Membrele șopârlei monitor
A - fata; B - înapoi:
1 - humerus, 2 - cubitus3 - raza, 4 - încheietura mâinii, 5 - metacarpul, 6 - falanga degetelor, 7 - articulația intercarpală, 8 - femurul, 9 - tibia, 10 - tibia, 11 - patella, 12 - tarsul 13 - articulația intertarsală, 14 - metatars
Membrele reptilelor sunt construite după modelul tipic al membrelor vertebrate terestre. Partea proximală a forelimb (fig. 89, A) este reprezentată de un os - humerus (humerus; fig. 89, 1), urmat de antebraț, care este format din două oase - ulnar (ulna; fig. 89, 2) și raza (raza; Fig. 89, 3). Încheietura (carpus, Fig. 89, 4) este formată din oase relativ mici, de obicei situate în două rânduri; în partea lor se află un alt os în formă de pere, luat pentru restul celui de-al șaselea deget. Metacarpul (metacarp; Fig. 89, 5) este alcătuit din cinci oase alungite de care sunt atașate falangele celor cinci degete (phalanges digitorum; Fig. 89, 6).
Ultimele falange poartă gheare. Articulația, care asigură mobilitatea mâinii, în reptile nu trece între oasele antebrațului și rândul proxim al oaselor încheieturii (ca în amfibieni), ci între rândurile proximale și distale ale oaselor încheietura mâinii. O astfel de îmbinare este numită intercarpală (Fig. 89, 7).
La nivelul membrului posterior se articulează elementul proximal - femurul (femur; Fig. 89, 8) articulația genunchiului cu o tibie formată din două oase de tibie - mari (tibia; fig. 89, 9) și mici (fibula; fig. 89, 10). Deasupra suprafeței frontale a acestei articulații se află un os mic - patella (fig. 89, 11). În tars (tarsus; Fig. 89, 12), rândul proximal al osiculelor fuzionează sau se conectează aproape nemișcat la oasele piciorului inferior, iar osiculele rândului distal sunt, de asemenea, strâns legate și parțial fuzionate cu oasele metatarsiene. Datorită acestui fapt, suprafața articulară de aici este situată nu între piciorul inferior și picior, ci între rândurile proximale și distale ale oaselor tarsale. O astfel de articulație este caracteristică reptilelor și se numește articulația intertarsală (Fig. 89, 13). Metatarsul (metatars; Fig. 89, 14) este format din cinci oase alungite de care sunt atașate falangele celor cinci degete (falanges digitorum; Fig. 89, 6). Falangele terminale poartă gheare.
CONCLUZIE
Reptile (Reptilia) - prima clasă a grupului de adevărate vertebrate terestre (Amniota), ai căror reprezentanți au un complex de adaptări pentru a trăi pe uscat fără nicio legătură cu mediul acvatic.
S-a spus mai sus că legătura strânsă dintre strămoșii reptilelor - amfibieni - cu corpurile de apă (sau cel puțin cu locuri cu umiditate ridicată) se datora în principal a două caracteristici biologice: tipul acvatic de reproducere și dualitatea sistemului respirator (plămâni și piele). Aceste caracteristici sunt reflectate la rândul lor asupra naturii metabolismului apei: trăind într-un mediu umed, amfibienii nu au mecanisme morfofiziologice pentru a reduce pierderea de umiditate a organismului. Dezvoltarea deplină a terenului este de neconceput fără transformări fundamentale precum reproducerea, respirația și metabolismul apei.
Implementarea acestor transformări caracterizează clasa reptilelor.
Cele mai specifice trăsături ale acestei clase (și alte amniote) ca vertebrate terestre primare sunt asociate cu adaptarea la viață în aerul atmosferic relativ uscat. Caracteristicile structurale specifice ale reptilelor sunt asociate în principal cu adaptarea la utilizarea mai economică a umidității. Pielea reptilelor este uscată și lipsită de glande (cu excepția unui număr mic de glande specializate care secretă un secret relativ gros). Straturile exterioare ale epidermei sunt reprezentate de celule moarte umplute cu materie coarnă (keratogialină).
Acest strat cornos protejează corpul de uscare și, de asemenea, îmbunătățește foarte mult protecția împotriva deteriorarii chimice și mecanice. Sub stratul cornos, stratul malpigian al epidermei este reprezentat de celulele vii. Pe măsură ce celulele moarte sunt uzate, acestea sunt înlocuite de celule din straturile inferioare, în a căror protoplasmă apar la început granule separate de keratogialină, care ulterior umplu întreaga celulă. În plus față de keratinizarea generală a stratului superior al epidermei, reptilele sunt caracterizate prin formarea în piele a diferitelor forme de solzi coarnă care acoperă suprafața pielii. Cantarul asigură o protecție mecanică a pielii și contribuie la o reducere și mai mare a pierderii de umiditate prin integument; unele reptile moderne (de exemplu, crocodili) au plăci osoase sub solzi excitat.
Toate aceste transformări, benefice din punct de vedere biologic în ceea ce privește menținerea unui echilibru de apă mai economic al organismului, au fost posibile în primul rând, deoarece pielea reptilelor nu participă la respirație, iar acest lucru se datorează dezvoltării progresive a sistemului respirator (vezi mai jos).
O importanță deosebită în evoluția vertebratelor terestre superioare este transformarea sistemului excretor. Spre deosebire de clasele studiate anterior, reptilele (ca și celelalte Amniota) au rinichi pelvini (metanefros). Spre deosebire de rinichii trunchiului (mesonephros), anamneza tubilor excretori ai rinichiului pelvin a pierdut complet contactul cu cavitatea corpului; produsele metabolice intră în tubii excretori numai din plexul capilarelor sanguine - glomerul malpighian. Este închis într-o ieșire scobită a capătului tubului - capsula Bowman. Acest lucru duce la faptul că în procesul de izolare a produselor de disimilare, organismul pierde mai puțină apă (lichidul cavității nu este excretat). Vasele de sânge care ies din glomerul malpighian formează oa doua rețea de capilare în jurul peretelui tubului excretor, apoi se scurg în vasele mai mari ale sistemului venos. Prin această a doua rețea de capilare, apa, carbohidrații și unele săruri sunt reabsorbite în sânge, astfel încât așa-numita „urină secundară” care este eliberată devine mai concentrată și are o compoziție diferită de „urina primară” formată în capsulele Bowman.
Rinichiul pelvin (metanefric) are propriul ureter, nu omolog la canalele Müller sau Wolf. Canalul Lupului se păstrează doar la bărbați, unde este conectat prin epididimă la glanda genitală și joacă rolul vaselor deferenți. Canalul Mueller la bărbați este redus. Femeile, pe de altă parte, păstrează doar canalele Müller; ele îndeplinesc funcția oviductelor și se deschid în cavitatea corpului cu găuri - pâlnii ale oviductului. Canalele de lup la femei sunt reduse.
Astfel, principalele caracteristici ale sistemului excretor al vertebratelor terestre superioare sunt pierderea comunicării între rinichi și cavitatea corpului și întărirea sistemului de absorbție inversă a apei din urina primară în sânge. Aceste proprietăți asigură implementarea funcției excretorii cu un consum minim de apă, ceea ce pentru locuitorii terenului, așa cum s-a menționat deja, are o semnificație biologică mare. La aceasta putem adăuga că, printre vertebratele terestre, intensitatea filtrării generale și absorbția inversă a apei este diferită la animalele care trăiesc în condiții diferite. Așadar, printre reptile, crocodilii și țestoasele (în mare parte forme de apă dulce) au glomeruli bine dezvoltați și secretă urină lichidă, în timp ce la șopârlele și șerpii, glomerulii sunt vizibil mai slabi, iar urina este solidă sau semi-lichidă.
Întregul complex al caracteristicilor luate în considerare, care oferă un echilibru hidric mai economic al reptilelor, stă la baza separării finale a vertebratelor terestre superioare de mediul acvatic și a populației lor răspândite din diferite biotopuri terestre.
Alături de aceste caracteristici, reptilele sunt caracterizate de o serie de transformări care au o valoare progresivă comună în evoluția vertebratelor.
Sistemul respirator s-a schimbat semnificativ. A apărut o trahee, a cărei lungime depinde de lungimea gâtului. Plămânii reptilelor se caracterizează printr-o structură internă mai complexă decât amfibienii: au traversele interne care le împart în departamente. De la aceste bare transversale, la rândul lor, se despart partiții, dezvoltate în grade diferite în diferite grupuri. Între sept, se formează căi respiratorii intrapulmonare, care servesc ca o continuare a bronhiilor și, în unele cazuri, sunt împărțite în bronhiile intrapulmonare de ordinul doi. Pereții interiori ai camerelor formate de acest întreg sistem de fascicule încrucișate au o structură celulară, astfel încât, în consecință, plămânii capătă o structură spongioasă complexă. Un astfel de dispozitiv al plămânilor crește semnificativ suprafața respiratorie totală și, în consecință, intensitatea respirației. Datorită acestor dispozitive, cererea de oxigen a organismului de reptile este complet asigurată doar de respirația pulmonară.
Este extrem de important ca actul de respirație la reptile să se realizeze prin extinderea și contractarea activă a cavității abdominale sub acțiunea mușchilor abdominali și costali. Acest tip de respirație asigură o ventilație mai intensă a plămânilor cu un volum mare de aer; aspectul său este asociat cu dezvoltarea unui piept mobil.
Transformări progresive semnificative apar în sistemul circulator al reptilelor (o schemă pentru separarea fluxurilor sanguine venoase și arteriale în inima reptilelor a fost dată în descrierea sistemului circulator).
Datorită apariției unui sept incomplet în ventricul și plecarea independentă din diferitele sale departamente ale principalelor trunchiuri arteriale (care este asociată cu o reducere a conului arterial), se realizează o separare destul de completă a fluxurilor de sânge arterial și venos. În momentul sistolei ventriculare, cavitățile sale interne sunt izolate de ceva timp; în timp ce sângele venos intră în artera pulmonară, sângele arterial intră în arcul aortic drept, iar amestecul venos arterial intră în arcul aortic stâng. Deoarece atât arterele carotide, cât și subclaviene provin din arcul aortic drept, capul și fața corpului sunt furnizate cu sânge arterial aproape pur (de exemplu, în țestoasele din arterele carotide, saturația de oxigen din sânge a depășit 90% în jumătate din cazuri). Navele care se extind sub conexiunea arcadelor aortice transportă sânge care conține o anumită impuritate venoasă către organele jumătății posterioare a corpului.
Septul orizontal care împarte ventriculul inimii reptile în două camere principale nu este omolog cu septul interventricular al păsărilor și mamiferelor. Dintre toate grupurile de reptile doar la crocodili, un nou - vertical - sept apare în ventricul, înlocuind orizontala antică. Acest sept vertical împarte complet ventriculul cardiac al crocodililor în două camere izolate: dreapta (venoasă) și stânga (arterială). Astfel, inima crocodililor poate fi numită în patru camere: are două atrii izolate unul de celălalt și două ventricule complet separate. Cu toate acestea, împreună cu separarea completă a ventriculelor, crocodilii păstrează ambele arcade aortice, care formează o cruce caracteristică tuturor reptilelor: dreapta începe în stânga și stânga în ventriculul drept. La intersecție există o așa-numită deschidere de pannici care leagă cavitățile ambelor arcade aortice.
Arterele carotide și subclaviene, ca toate reptilele, se extind din arcul aortic drept. Arcul aortic stâng, care înconjoară inima, dă vase de sânge către intestin și, printr-o conductă subțire, se conectează la arcul aortic drept, dând naștere arterei spinării. Conform unor rapoarte, la baza arcului aortic stâng se găsesc valve și un sistem de cartilaj care împiedică intrarea sângelui venos în el cu sistolă ventriculară.
Unii autori explică păstrarea atât a arcadelor aortice, cât și a conexiunii lor la locul de intersecție cu caracteristicile ecologice ale crocodililor: în timp ce sub apă, circulația pulmonară circulă puțin sânge, deoarece respirația este suspendată, iar excesul de sânge din circulația pulmonară trece într-un cerc mare, susținând astfel vasele de sânge tensiunea arterială normală. Este posibil ca conservarea conductelor botalale (vasele care leagă arcadele aortice cu arterele pulmonare) la țestoase care duc un stil de viață acvatic să fie de asemenea la fel de importantă. Ca și canalele carotide (conexiunea arcadelor aortice cu arterele carotide) ale șerpilor și șopârlelor, aceste formațiuni sunt rudimentele arcadelor arteriale ale reptilelor ancestrale îndepărtate, care au fost reduse la respirația pulmonară. Cea mai primitivă dintre reptilele vii, hatteria, are atât conducte carotide cât și ductale.
central sistemul nervos reptilele se caracterizează printr-o creștere a dimensiunii relative a creierului și dezvoltarea progresivă a emisferelor anterioare, în acoperișul căruia apare prima dată materia cenușie - cortexul cerebral (încă slab dezvoltat).
Printre trăsăturile progresive ale scheletului reptilelor este importantă apariția unui piept închis. În plus față de valoarea deja indicată a acestei transformări în intensificarea respirației, pieptul asigură conectarea brâului de umăr cu scheletul axial și, prin urmare, fixarea ei mai fermă, ceea ce duce la creșterea funcția de sprijin membrelor. La șerpii care și-au pierdut membrele împerecheate pentru a doua oară, centurile le sunt reduse.
Datorită singurului condil occipital, structura specifică a primelor vertebre cervicale, care permite atlasului să se rotească în jurul procesului asemănător cu dinții epistrofiei și apariția multor vertebre cervicale care sunt articulate mobil între ele, mobilitatea capului reptilei a crescut semnificativ. Aceasta are o importanță biologică deosebită în orientarea, găsirea și obținerea de alimente etc.
Modificările evolutive ale acoperișului craniului sunt foarte semnificative. În cele mai vechi reptile, pereții superiori și laterali ai craniului erau compuși din numeroase oase integumentare și aveau aspectul unui carapace osos solid. Un astfel de craniu se numea matlasat. O evoluție ulterioară a mers pe calea reducerii treptate a oaselor integumentare ale craniului matlasat, astfel încât depresiunile (gropi temporale) s-au format în locul oaselor reduse, delimitate de poduri osoase relativ subțiri (arcade temporale). Aceste transformări ale craniului, pe de o parte, au dus la relieful său, care este important atunci când trăim într-un mediu de aer mult mai puțin dens decât cel acvatic; pe de altă parte, au fost utilizate depresiuni formate la locul oaselor reduse ale acoperișului craniului. Pentru a se acomoda cu mușchii maxilarului. Dezvoltarea mușchilor maxilarului este direct legată de o creștere a activității nutriționale.
În majoritatea grupurilor de reptile în timpul reducerii acoperișului secundar al craniului, s-au format două fosele temporale: partea superioară, delimitată extern de arcul temporal superior (oasele frontale și scaltoase posterioare), iar cea laterală, delimitată deasupra superioară și în afara oaselor inferioare (oasele zigomatice și pătrat-zigomatic). Reptilele cu acest tip de craniu aparțin grupului diapsid (bipedal). La unii reprezentanți ai acestui grup, reducerea oaselor a mers mai departe în direcția pierderii unuia sau chiar a ambelor arcade temporale. Un craniu diapsid tipic pronunțat este caracteristic formelor vii pentru hatteria și crocodili. În multe șopârlele, arcul temporal inferior este parțial redus, iar în unele, cel superior. Ambele arcade temporale sunt reduse la șerpi. Păsările au evoluat din reptile diapside (la aceste animale arcul temporal inferior a fost păstrat complet, dar cel superior a fost redus).
În unele reptile, evoluția regiunii temporale a craniului a mers pe calea formării unei singure fose temporale, limitată la un singur arc temporal compus din oasele zigomatice și scaltoase. Întrucât osul zigomatic este un element caracteristic al inferiorului, iar osul scalz este arcadele temporale superioare ale craniului diapsid, un astfel de arc se numește mixt, iar reptilele cu o astfel de structură a craniului aparțin grupului sinapidelor (reptile cu un arc mixt). Printre reptilele moderne, nu există forme de sinapsă. Acest tip de craniu este caracteristic descendenților reptilelor sinapidelor - mamifere.
La una dintre ramurile reptilelor primitive, reducerea oaselor integumentare ale acoperișului craniului a mers altfel: formarea unei crestături în partea occipitală a craniului integumentar a dus la formarea de fiecare parte a unei cavități - o falsă fosa temporală, acoperită la exterior cu o punte osoasă largă a oaselor posterioare, solzoase și pătrat-zigomate. Astfel de reptile aparțin grupului anapsidului (nejustificat). Dintre reptilele vii, țestoasele aparțin acestui grup.
Setul de caracteristici progresive ale reptilelor a dus la o creștere generală a energiei vitale a activității acestor animale și la apariția dintre ele a unei largi varietăți de forme de viață, reflectând adaptările la diverse condiții de viață. Spre deosebire de amfibieni, consumând exclusiv hrană pentru animale, printre reptile există forme erbivore (cum ar fi, de exemplu, țestoasele terestre). Dar totuși, majoritatea reptilelor moderne sunt carnivorele. O mobilitate mai mare în comparație cu amfibienii le permite să vâneze prada mobilă, ajungând uneori la dimensiuni semnificative. Într-o serie de cazuri, succesul vânătorii este facilitat de o serie de dispozitive speciale: dinți otrăvitori la unele specii de șerpi, culoarea protectoare caracteristică multor reptile etc.
Majoritatea reptilelor sunt animale terestre. Pe uscat, diferite tipuri de reptile se găsesc într-o mare varietate de condiții: în deșerturi, păduri de vegetație mlăștinoasă, păduri, printre roci etc. stați pe suprafețe abrupte) pentru a evita atacurile prădătorilor (îngroparea în nisip, autotomia cozii de coadă etc.) etc.
Multe reptile au trecut din nou la viața acvatică (țestoase de mare și apă dulce, crocodili, șerpi acvatici). Unele dintre aceste specii își petrec aproape întreaga viață în apă, dar merg neapărat pe pământ pentru reproducere. Șerpii de mare au dezvoltat abilitatea de a ovipari, ceea ce le-a permis să treacă din nou pe deplin la modul de viață acvatic (printre reptilele dispărute, ictiosaurii, care dețin și capacitatea de a ovipari), au dus un mod de viață complet acvatic).
Reptilele au atins cea mai mare diversitate de forme de viață în timpul perioadei evolutive a acestei clase - în epoca mezozoică. Printre reptilele fosile, sunt cunoscute ierbivorele și carnivorele (diferite grupuri de dinozauri), locuitori profund specializați ai mediului acvatic (ictiosauri) și forme zburătoare (pterodactile, ramphorinchs), precum și multe alte grupuri biologice. Răcirea generală a climei care a venit după Mesozoic nu a contribuit la prosperitatea suplimentară a reptilelor: la fel ca amfibienii, reptilele sunt poikilotermice și nu pot ține o temperatură ridicată a corpului pentru o perioadă lungă de timp în timpul apariției reci.
Majoritatea speciilor de reptile moderne trăiesc în climă caldă; speciile obișnuite în zona temperată petrec o parte semnificativă a anului într-o amețire. În aceste condiții, avantajele evidente s-au dovedit a fi „descendenții” lor - păsări și mamifere, în evoluția cărora a apărut o nouă calitate valoroasă - homoyotherm. Capacitatea de menținere a vieții active într-o gamă largă de temperaturi ambientale este unul dintre motivele principale pentru determinarea poziției predominante a păsărilor și mamiferelor în fauna modernă a vertebratelor.
Citirea ulterioară
Gurtovoi N.N., Matveev B.S. Dzerzhinsky F. Ya. Zootomie practică a vertebratelor. Amfibieni, reptile. M., 1978
Herpetologie Terentyev P. V. M., 1961.
Schmalhausen I. I. Fundamentele anatomiei comparative a vertebratelor. M., 1947
Coloana vertebrală combină 4 secții: col uterin, lombo-toracic, sacral și caudal. Corpurile vertebrale sunt concave în față și convexe în spate - vertebre întregi.
Arcadele superioare se termină cu un proces spinos bine definit; o pereche de procese articulare anterioare scurte se îndepărtează de marginea anterioară a arcului superior și o pereche de procese articulare posterioare se îndepărtează de marginea posterioară a aceluiași arc. Nu există semne de reziduuri de coarde la adulți.
Schelet de șopârlă (conform lui Bram):
1 - claviculă, 2 - omoplat, 3 - umăr, 4 - rază, 5 - ulna, 6 - încheietură, 7 - încheietura mâinii, 8 - falangă a degetelor, 9 - coaste, 10 - pelvis, 11 - coapsa, 12 - mare tibia, 13 - tibia, 14 - tarsus, 15 - metatarsus, 16 - vertebre caudale
coloanei vertebrale de col uterin reprezentate de opt vertebre. Partea din față a acestora, numită atlas, sau atlas, are o formă inelară caracteristică tuturor amniotelor. Pe partea frontală a părții sale inferioare se află o suprafață articulară nepereche, cu ajutorul căreia vertebra se articulează cu craniul. Gaura atlasului este împărțită de un pachet în două jumătăți - superior și inferior. Măduva spinării trece prin orificiul superior, iar procesul dentoid (procese odontoideus) intră în gaura inferioară, care este ferm conectată, dar nu este fuzionată la a doua vertebră, numită epistrofă (epistrofă). După cum dovedesc datele embriologice, dentatul este corpul detașat al primei vertebre. Astfel, atlasul este mobil în raport cu propriul corp. Ultimele trei vertebre cervicale poartă o pereche de coaste cervicale bine dezvoltate, care sunt atașate mobil de corpurile vertebrale, cresc treptat posterior și se termină liber, fără a ajunge la stern. În general, partea cervicală a reptilelor diferă de cea a amfibienilor nu numai într-un număr mare de vertebre, ci și într-una mai mobilă: articularea primei vertebre atât cu craniul, cât și cu a doua vertebră. Drept urmare, capul primește o libertate de mișcare mult mai mare.
coloanei vertebrale lombo-toracice șopârlă conține 22 vertebre. Toate sunt echipate cu coaste mobile, a căror lungime scade treptat spre spate. Fiecare coaste constă din cartilajul osos superior și inferior. Primele cinci vertebre lombare-toracice (de fapt vertebrele toracice) au coaste deosebit de lungi care se atașează de stern, formând torace (Thorax). Sternul (sternul) este cartilaginos și are forma unei plăci rombice. Dezvoltarea embrionară a sternului amniotului arată că este formată prin intergroparea distaluluicapătul coastelor și, prin urmare, nu este omolog cu „sternul” amfibienilor, format independent. Astfel, în șopârlă, este deja conturată o diviziune a trunchiului coloanei vertebrale în coloana toracică și lombară, extrem de caracteristică pentru amnioti.
Secțiune de coadă este format din numeroase vertebre (câteva zeci). Cele anterioare sunt similare cu cele sacre, dar corpurile lor sunt mai lungi, procesele transversale sunt mai subțiri, iar procesele spinoase sunt mai lungi. Spre spate, vertebrele caudale scad treptat în dimensiuni, își pierd procesele și se transformă în mici oase în formă de tijă.
Articole mai interesante
Dezvoltarea și complicația sistemului motor al reptilelor este cauzată de trecerea la un habitat predominant pe uscat și de necesitatea depășirii denivelărilor reliefului său. Principalele modificări ale aparatului de susținere a reptilelor, în comparație cu amfibienii, sunt asociate cu mobilitate sporitănecesare pentru căutarea cu succes a hranei, a reproducerii, a protecției împotriva pericolului etc. Acest lucru este reflectat în întărirea craniului, o mai mare diferențiere a scheletului axial, întărirea membrelor și a centurilor, dezvoltarea în continuare a mușchilor specializați.
Scheletul reptilelor este caracterizat prin dezvoltare semnificativă a osului elemente și conexe reducerea cartilajelor formațiuni. Un număr deosebit de mare de osificări integumentare compune acoperișul, părțile laterale și partea inferioară a craniului creierului. Schimbări semnificative și departamentul visceral. Aparatul maxilarului a devenit mai masiv, oasele maxilarului superior sunt adânc înrădăcinate partea de sus departamentul creierului. În același timp, craniul a dobândit mobilitate în conexiunea secțiunilor cerebrale și viscerale. Deci, șerpii partea inferioară craniul este un complex sistem articulat cu pârghie, care vă permite să înghițiți prada întreagă mare (boas, șerpi de ou etc.). Contribuie la asta gropi și arcade temporale, fiind un loc de atașament al unui puternic și mușchii elastici. Scheletul capului se articulează cu coloana vertebrală prin unic condilformat din oasele gâtului. În general, craniul a devenit mai puternic și, în același timp, mai ușor.
Complicația diferențierii coloanei vertebrale se manifestă printr-o creștere a numărului de vertebre în secțiunile trunchiului. De o importanță deosebită marirea colului uterin departament, care determină mobilitatea capului, care, la rândul său, îmbunătățește orientarea pe sol și ajută la accelerarea mișcării. Fuzionând partea din față a coastelor cu sternul, se formează torace. Se dezvoltă în zona pieptului mușchii intercostali, care determină mecanismul respirator la multe specii.
extremitate majoritatea reptilelor sunt scurte, atașate de centurile membrelor și au mușchii specializați. În zona de conectare a antebrațului cu peria se formează articulația intercarpală. O formațiune similară are loc la membrele posterioare, unde în partea de gleznă se formează articulația intertarsală. Aceste formațiuni contribuie în mod semnificativ la consolidarea și mobilitatea extremităților inferioare. Curea membrelor păstrând o cantitate semnificativă de cartilaj, membrele sunt atașate la schelet axial.
Reptilele care și-au pierdut membrele, centurile și pieptul (șerpi, șopârlele fără pereche), au coaste lungi, trunchi puternic și mușchi subcutanat. Într-o serie de reptile (țestoase), pierderea vitezei de mișcare este asociată cu dezvoltarea unui dispozitiv special de protecție - coajă fixămișcare de prunc. Caracteristicile structurale ale scheletului sunt și ele specii acvatice secundare (broaște țestoase marine, șerpi de mare, iguane, crocodili).
După ce vă familiarizați cu caracteristicile de bază ale scheletului, putem trece la o examinare mai detaliată a structurii părților sale constitutive.
craniu
Craniul este alungit, cu un conținut semnificativ de elemente osoase (Fig. 51). O formă particulară a craniului, raportul dintre mărimile componentelor sale în diferite reptile este determinat tipul de putere și metoda de captare produse alimentare.
Craniul creierului întărită prin creșterea numărului de oase și fuziunea lor solidă între ele.
Acoperișul cranii formate din pereche oasele nazale, anterioare frontale, frontale, posterioare (postorbitale) și parietale. fără egal osul interparietal are foramen parietal autoritate. Spre laturile acestor elemente cresc oase lacrimale și infraorbitarecare încadrează partea superioară a orbitei ochilor.
fund se formează cranii osul sfenoid principal cu rudimentare parasphenoid și deschizători gemenepe laturile cărora se află choanae. Elementele secțiunii viscerale iau parte și la formarea fundului craniului creierului. Oase pterygoid articula cu pătrat, care la șerpi și șopârlele sunt conectate mobil la craniul creierului și acționează ca o suspensie pentru aparatul maxilar. între pterygoids și maxilareste situat osul transvers, care în plus consolidează craniul.
occipital este format din patru oase - lateral, supraoccipital și pereche lateral. Ca urmare a creșterii lor, un condil occipital. În partea de jos a departamentului se află foramen occipital pentru a conecta creierul și măduva spinării.
Partea laterală craniile alcătuiesc trei perechi oase auriculare (anteroposterior, auricular superior și auricular posterior), care sunt implicați în educație gropi temporale.Oasele superioare și posterioare sunt contopite cu oasele regiunii occipitale. Lângă ele sunt temporal, scarăpărți superioare în formă de aripă oase. împerecheat oasele coloanei conectați acoperișul craniului la partea inferioară. Socluri mari pentru ochi formează elemente osoase adiacente - infraorbital, postorbital, zigomatic, lacrimal. Septul interorbital format din depășirile oaselor frontale și parietale. În fața craniului creierului oasele maxilare și intermaxilare se mărunțesc. olfactivzona craniului cartilaj.
Fig. 51. Craniul unei șopârle deasupra (A), de sub (B), laterală (C) și a maxilarului său inferior:
1 - foramenul occipital, 2 - osul occipital lateral, 3 - osul occipital superior, 4 - osul occipital principal, 5 - condilul occipital, 6 - osul sfenoid principal, 7 - vomerul, 8 - choans, 9 - parafenoidul vestigial, 10 - parietal os, 11 - o deschidere pentru ochiul parietal, 12 - osul frontal, 13 - osul nazal, 14 - osul maxilar, 15 - osul maxilar, 16 - osul prefrontal, 17 - osul lacrimal, 18 - osul infraorbital, 19 - osul orbital, 20 - os scalp, 21 - osul temporal, 22 - os zigomatic, 23 - nară, 24 - os pătrat, 25 - os palatin, 26 - os pterigoid, 27 - os coloană, 28 - os transvers, 29 - os articular, 30 - os dentar, 31 - os unghiular, 32 - osul arcuit, 33 - proces coronoid
Craniul unui număr de reptile este caracterizat prin prezența unor bine definite arcade temporale (sunt reduse în șopârlă). Numai la șopârlă monitor arcade temporale superioarela educația căreia iau parte orbitala si solzoasa oase. Aceste arcuri delimitează lateral gropi temporale. Crocodilii, împreună cu arcurile superioare, au inferiorcare se formează maxilar și zigomaticoseminte (craniu diapsid) (Fig. 52). în broască țestoasănu există gropi temporale și arcade temporale ( craniu de tip anapsid).
Craniu visceralconstă din maxilar și aparat hioid . osemintepartea de sus falcă (intermaxilar și maxilar), așa cum sa menționat deja, au fost întărite și adânc înrădăcinate în craniul creierului. Are loc autostiliya - conexiunea directă a craniului visceral cu creierul. Ca urmare a osificării și transformării cartilajului pătrat-pătrat pătrat și columnar oase. Palatin și pterygoid elementele sunt de origine cutanată și sunt implicate în formarea fundului craniului creierului.
O serie de reptile (crocodili, țestoase) au un loc palatul osos secundar, care a fost format prin creșterea unui număr de oase - intermaxilar, maxilar, palatin și în formă de aripă. Crocodilii s-au schimbat și poziția nării: nările exterioare sunt ridicate deasupra suprafeței capului, iar coana este deplasată în regiunea occipitală a craniului. Aceste transformări stau la baza separării nazofaringelui de cavitatea bucală, ceea ce permite animalelor să-și țină gura deschisă sub apă și, în același timp, să respire prin nările expuse la suprafața apei.
Maxilarul inferior format prin osificare cartilajul lui Meckel și apariții articulat, dentar, unghiular, arcuit, coronal oase și o serie de formațiuni mai mici. Dintre oasele enumerate, numai articularul are origine chondrală, toate celelalte sunt integumentare.
Aparat sublingual reprezintă record (copula omolog) cu trei perechi de coarne - derivatele hioidelor și arcadelor ramurale. Hyomandibular, ca și în amfibieni, sub formă de osic auditiv ( etrieri)situat în urechea medie.
Echipamente și materiale
1. Scheletele unei șopârlă, țestoase, șerpi (un set pentru doi elevi).
2. Vârfuri de șopârlă din diferite părți ale corpului.
3. Membrele anterioare și posterioare ale șopârlei cu centuri.
4. Craniul unei șopârle, țestoase, șerpi.
5. Tabelele: 1) scheletul unei șopârle; 2) structura vertebrelor din diferite părți ale corpului; 3) craniu (vedere de sus și de jos); 4) scheletul membrelor și al centurilor.
Observații introductive
Principala direcție a evoluției reptilelor a fost îmbunătățirea aparatului locomotor și creșterea diversității mișcărilor. Reptilele (moderne și dispărute) demonstrează toate principalele moduri de mișcare cunoscute de vertebrate.
Modul de mișcare evolutiv original și tipic a fost păstrat în șopârlele moderne. La fel ca în cazul amfibienilor cu coada, mișcarea se realizează prin rearanjarea alternativă a membrelor relativ scurte așezate în laterale
când aplecați corpul spre dreapta - spre stânga. Structura articulațiilor membrelor și modelul general de locomoție în șopârlele moderne sunt modificate doar ușor în comparație cu reptilele timpurii.
Eficacitatea acestei metode de mișcare printre reptile a devenit mai mare în comparație cu amfibienii. Acest lucru s-a realizat, pe de o parte, prin întărirea și diferențierea mușchilor corpului și membrelor, iar pe de altă parte, prin diferențierea scheletului și consolidarea și transformarea nodurilor și articulațiilor individuale ale acestuia. Consolidarea mușchilor membrelor, dezvoltarea mușchilor longitudinali ai corpului, deplasarea într-o măsură sau alta a membrelor de sub corp - toate acestea au făcut posibilă mișcarea rapidă cu corpul ridicându-se deasupra solului (alergare). Consolidarea mușchilor și mișcarea într-un mediu divers a fost însoțită de o creștere a sarcinii pe schelet. Aceasta a necesitat consolidarea sa. Prin urmare, țesătură de fibră grosieră oase tubulare schimbat în fibroase fine, claviculele au început să joace rolul unui fel de distanțier între părțile dorsale (marginea distală a scapulei) și părțile ventrale (sternului) ale brâului de umăr. Cel mai frecvent a fost tipul procentual de vertebre. Toate vertebrele trunchiului, precum și o parte a colului cervical și caudal, poartă coastele mobile. Mișcări intense de flexie - extensia mâinii și piciorului a devenit posibilă datorită apariției articulațiilor intercarpale și intertarice.
Reptilele sunt caracterizate printr-o mobilitate mai mare a capului. Aceasta este asigurată de articularea extrem de mobilă a craniului cu coloanei cervicale coloana vertebrală: în primul rând, craniul se conectează la prima vertebră cervicală cu un singur condil; în al doilea rând, primele două vertebre cervicale (atlas și epistrofie) sunt modificate astfel încât mișcările capului în jurul axei longitudinale a corpului sunt posibile. Mobilitatea capului și a gâtului, combinată cu capacitatea de a se deplasa rapid în jurul substratului, a oferit reptilelor posibilități largi pentru găsirea hranei. Compresia puternică a fălcilor s-a dovedit a fi importantă pentru capturarea pradelor diverse. În acest sens, s-au dezvoltat mușchi de mestecat puternici și au fost reconstruite acoperișul și laturile craniului - au apărut gropi temporale, ceea ce a mărit semnificativ suprafața pentru atașarea mușchilor maxilarului. Deformarea craniului sub compresie mesteca muschii împiedicat de oasele coloanei.
Legătura coastelor cu sternul și formarea toracelui în acest fel este o altă transformare majoră a scheletului reptilelor. Drept urmare, rata de ventilație a plămânilor a crescut brusc și a crescut volumul de aer pompat. Aceasta a contribuit la respingerea respirației pielii și la creșterea intensității schimbului de gaze.
În general, scheletul reptilelor poartă un complex de trăsături care demonstrează un nivel mai ridicat de organizare a acestor animale și adaptări mai profunde la viața pe uscat.
Explorați scheletul reptilelor.
ia în considerare:
craniu
Craniul creierului
occipital: top; principal și două oase laterale occipitale; foramen occipital; condil occipital.
Partile craniului: malar; solzos; webbed; bar; pătrat; oase de ureche.
Acoperiș craniu: nazal; prefrontal; frontală; parietal; interparietal; rupe; fruntea; oasele postorbitale.
Fundul craniului: osul sfenoid principal; deschizător; rudimentele palatului osos secundar.
Craniu visceral
Maxilarul superior: maxilar; oase intermaxilare.
Maxilarul inferior: dentare; articular; unghi; oase arcuite;
Aparate sublinguale: os hioid cu două perechi de coarne.
coloană vertebrală
Coloana vertebrală: gât; sân; lombare; sacral; coadă; vertebra trunchiului; Atlanta; epistrophe. piept: ster; coaste.
Curea membrelor
Brâu de umăr: ster; apucător de piept; coracoid; omoplat; clavicula.
Brâu pelvin: iliacă; sciatic; oase pubiene; acetabulum.
Membre pereche
a membrelor anterioare: umăr; antebraț (ulna și raza); încheietura mâinii; metacarp; falangele degetelor; articulația intercarpală.
Membrul posterior: coapsa; tambur (tibia și tibia); Tars; metatars; falangele degetelor; articulația intearsală.
schiţa:
1) craniu (vedere de sus, vedere laterală, de jos); 2) centura primelor; 3) brâu pelvin și coloana vertebrală sacrală.
Structura scheletului
Craniul creierului
În șopârlele, craniul este aproape complet osificat și constă dintr-un număr mare de oase, formând un loc destul de masiv cutie de creier (Fig. 70, 71).
Regiunea occipitală a craniului este format din patru oase occipitale, limitând foramenul occipital. Osul occipital principal împreună cu două oase laterale occipitale formează condilul occipital, care intră în fosa articulară a atlasului. Osul occipital superior sub forma unui arc acoperă foramenul occipital de sus și intră în spatele acoperișului craniului.
Partile craniului în zona capsulei auditive sunt formate din trei oase ale urechii, care sunt implicate în formarea gropilor temporale.
Fig. 70. Structura craniului unei șopârle:
A - vedere de sus; B - vedere de jos; - vedere laterală; 1
- foramen occipital; 2
- os occipital lateral; 3
- os occipital superior; 4
- osul occipital principal; 5
- condil occipital; 6
- principalul os sfenoid; 7
- deschizator; 8
- choans; 9
- parafenoid; 10
- os parietal; 11
- gaură pentru organul parietal; 12
- os frontal; 13
- os nazal; 14
- os intermaxilar; 15
- osul maxilar; 16
- osul prefrontal; 17
- os lacrimal; 18
- oase infraorbitare; 19
- os postorbital (posterior); 20
- osul scalcios; 21
- os temporal; 22
- os zigomatic; 23
- nara; 24
- os pătrat; 25
- os palatin; 26
- os pterygoid; 27
- osul coloanei; 28
- osul transvers; 29
- os articular; 30
- osul dintelui; 31
- os unghiular; 32
- osul arcuit; 33
- os coronarian

Fig. 71. Structura craniului unui șopârlă monitor:
A - vedere de sus B - vedere de jos 1
- os intermaxilar, 2
- os nazal 3
- osul prefrontal, 4
- os frontal 5
- os lacrimal 6
- os infraorbital 7
- os parietal, 8
- os occipital superior, 9
- osul occipital principal, 10
- os occipital lateral, 11
- osul solut 12
- os pătrat 13
- os postorbital (posterior), 14
- os zigomatic, 75 - os maxilar, 16
- deschizător 17
- os palatin, 18
- osul transvers 19
- osul pterigoid 20
- osul sfenoid principal, 21
- foramen occipital; 22
- os zigomatic pătrat; 23
- condil occipital
Oasele anterioare sunt bine dezvoltate și izolate, oasele aurale superioare fuzionează cu osul occipital superior, iar oasele posterioare cu oasele occipitale laterale. Pe părțile laterale ale craniului sunt prize mari pentru ochi. Septul interorbital este format prin procese ale oaselor frontale și parietale ale acoperișului craniului. În spatele orbitei se află oasele postorbite, zigomatice și scaltoase. Oasele lacrimale mici sunt situate în colțul frontal al orbitei, iar o serie de oase infraorbite mici mărginesc orbita de sus. Partea anterioară a craniului este formată din derivați ai craniului visceral - oasele maxilare și intermaxilare. În zona olfactivă, șopârlă nu are osificare, capsula olfactivă cu nările exterioare este formată prin cartilaj.
Acoperiș craniu se formează oase nazale, prefrontal, frontale și parietale. Între oasele parietale se află un os interparietal nepereche cu o deschidere pentru organul parietal.
Fundul craniului format din principal os sfenoidcu care se fuzionează parafenoidul rudimentar. În partea din față a parafenoidului se află oasele palatine și deschiderile, pe laturile cărora
sunt hoanele. Unele oase ale craniului visceral sunt de asemenea implicate în formarea fundului cutiei creierului - acestea sunt palatine și oase pterygoid. Oasele pterygoid curbate cu capetele frontale se prind de oasele palatine, iar capetele posterioare se articulează cu sfenoidul principal și cu oasele pătrate.
În regiunea temporală a șopârlelor, fosa temporală este localizată, limitată de un singur arc temporal superior, care este format din oasele postorbitale și scaltoase. Arcul temporal inferior în șopârlele este redus. În profunzimea fosei temporale, oasele coloanei (în formă de elytra) sunt clar vizibile sub formă de mici strunguri care vin din oasele pterigoid și parietal.
Craniu visceral
Arcul falcului în șopârlele este puternic modificat. Ca urmare a osificării cartilajului pătrat-palatin, acestea au format oasele pătrate, pterigiene, coloanare (aripioare elitre) și palatine. Osul pătrat cu capătul său superior este articulat mobil cu un os solzos, iar cel inferior cu maxilarul inferior și servește ca suspensie pentru acesta. Apariția unei articulații între osul scalos și capătul superior al osului pătrat ne-a permis să creștem forța de compresiune a fălcilor și, se pare, a provocat succesul evolutiv al șopârlelor. Maxilarul superior format din oasele maxilare și intermaxilare pe care se află dinții. Între oasele pterygoid și maxilar se află oasele transversale, care acționează ca distanțiere și întăresc craniul.
Baza pentru dezvoltarea oaselor maxilarul inferior servește cartilaj meckel. Este înlocuit cu un os articular care se articulează cu un os pătrat. În plus, maxilarul inferior include o serie de oase integumentare: cele mai mari oase dentare care transportă dinții, iar cele mai mici - coronare, unghiulare și arcuite.
Departamentul superior arcul sublingual - Hyomandibular - face parte din urechea medie, sub forma unui osic auditiv - un pas. Restul arcului hioid și rămășițele arcadelor branhiale sunt transformate în aparatul hioid, reprezentat de placa hioidă cartilaginoasă și trei perechi de coarne.
coloană vertebrală
Coloana vertebrală a reptilelor este împărțită în patru secțiuni: col uterin, toracic, lombar, sacral și caudal.
Majoritatea reptilelor au vertebre protselnye, adică anterioare concave. Corpul vertebral este alungit, este concavă în față,

Fig. 72. Primele două vertebre cervicale ale unei șopârle monitor:
A - atlas; B - epistrofie; 1
- arc superior; 2
- canalul spinal; 3
4
- proces epistrofie asemănător cu dinții; 5
- corp vertebral
duta superioară se încheie cu procesul spinos superior. Procesele articulare anterioare și posterioare se extind din corpul vertebral. La baza arcului superior se află suprafețe articulate pentru fixarea nervurilor.
Primele două vertebre cervicale sunt foarte diferite de restul. Prima vertebră cervicală - atlas - are forma unui inel împărțit de un pachet dens în jumătățile superioare și inferioare (Fig. 72). Pe suprafața frontală a atlasului există o fosă articulară pentru condilul occipital. Prin orificiul superior, creierul se conectează la măduva spinării, iar procesul asemănător cu al dinților vertebrei cervicale intră în gaura inferioară din spate - epistrofeya. Această articulare a craniului cu gâtul asigură mișcări complexe ale capului. Vertebrele cervicale posterioare poartă coaste.
La vertebre toracice ia parte coastecare se articulează cu sternul și formează cusca de coaste (coaste adevărate). Vertebrele lombare au și nervuri care nu se conectează la stern (coaste false). Două vertebre sacrale au procese transversale masive care se articulează cu brâul pelvin (vezi Fig. 75). Numărul vertebrelor caudale la șopârlele variază, ajungând la câteva zeci. Vertebrele caudale anterioare poartă procese transversale și spinoase (Fig. 73), iar la sfârșitul cozii, vertebrele sunt transformate în oase în formă de tijă. Corpurile aproape toate vertebrele caudale sunt separate printr-un strat subțire cartilaginos, de-a lungul căruia se produce o ruptură în timpul autotomiei.

Fig. 73. Structura vertebrei caudale anterioare a șopârlei:
A - vedere de sus; B - vedere laterală; 1
- corpul vertebrei; 2
- proces spinos superior; 3
- proces articular anterior; 4
- proces articular posterior; 5
- proces transversal; 6
- proces spinos inferior; 7
- arc inferior
Curea membrelor
Curea anterioară în șopârlele (Fig. 74) este în general asemănat similar cu brâul de umăr al amfibienilor, cu toate acestea, în comparație cu acesta, osificarea este foarte dezvoltată aici. La joncțiunea coracoidului cu scapula există o cavitate articulară pentru articularea cu humerusului. Cartilajul suprascapular crește până la marginea dorsală a scapulei, iar cartilajul procoracoid crește în fața coracoidului. Coracoidul și procoracoidul se îmbină, formând un singur element scheletal care este articulat cu sternul. În această articulație, sunt posibile mișcări de alunecare într-un plan orizontal de-a lungul marginii frontale a sternului, ceea ce crește lungimea pasului. De jos până la stern, crește un suprastern cruciform de origine integumentară. Claviculele leagă marginea dorsală a scapulei cu pectoralul. Astfel, spre deosebire de amfibieni, gulerele reptile sunt orientate pe un plan vertical și fac mai rezistentă brâul de umăr.
Brâu pelvin în șopârlele este bine dezvoltat și este format din două oase anonime, care au fost formate prin fuziunea oaselor pelvine - iliacă, sciatică și pubiană (Fig. 75). Există cavități articulare pentru capul femural (acetabulum). Oasele pubiene și ischiale dreapta și stânga sunt conectate prin simfiza cartilaginoasă. Brâul pelvin este ferm conectat la coloana vertebrală prin creșterea ileului la procesele transversale ale vertebrelor sacrale.
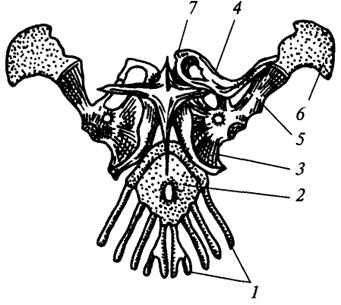
Fig. 74. Brâul de umăr al unei șopârle:
1
- primele patru coaste toracice; 2
- stern; 3
- coracoid; 4
- claviculă; 5
- scapula; 6
- cartilaj suprascapular; 7
- curea de piept

Fig. 75. Brâu pelvin al unei șopârle: br 1
- cavitatea articulară (acetabulară) pentru capul femural; 2
- ilium; 3
- os pubian; 4
- os ischial; 5
- simfiză
Membre pereche
Membrele reptilelor corespund structurii generale a membrelor cu cinci degete ale vertebratelor terestre. schelet a membrelor anterioare Acesta include humerusului, oase ulnare și radiale, două rânduri de oase mici ale încheieturii, un rând de oase metacarpiene și un rând de oase ale falangelor degetelor. schelet membrul posterior include femurul, tibia și tibia, două rânduri de oase tarsale mici, un număr de oase metatarsiene și o serie de falangele degetelor. Particularitățile structurii extremităților includ formarea interkarpalnogo (în prealabil) și tertarzalnogo invariante (în membrul posterior) al articulațiilor (fig. 76).

Fig. 76. Structura articulației intertarice a șopârlei (piciorul drept al șopârlei rapide de sus):
1
- oase tarsale proximale condensate; 2
- oasele distale ale tarsului; 3
- tibia; 4
- tibia mică; 5
- oase metatarsiene
Articulația intercarpală se formează între două rânduri de semințe de încheietura mâinii, articulația intertarsală se află între două rânduri de oase tarsale. Aceste articulații oferă posibilitatea unei flexiuni și extinderi puternice a mâinii și piciorului, ceea ce este important cu un tip particular de locomoție.