Koľko sakrálnych stavcov má jašterica. Muskuloskeletálny systém plazov všeobecná charakteristika kostry
TÉMA 14. SKELETON REBUMAN
SYSTÉMOVÉ POSTAVENIE CIEĽA
Podtyp Vertebrata, Vertebrata
Reptile Class, Reptilia
1. Squad Scale, Squamata
Zástupca - Varan Gray, Varanus griseus Daud.
2. Squad krokodíly, krokodíl
Zástupca - Mississippi Alligator, Alligator mississippiensts Daud.
3. Turtle Squad, Chelonia (Testudines)
Zástupca - korytnačina močiarna, okrúhly etnys je L.
MATERIÁL A ZARIADENIA
Namontované kostry jašteríc monitorov, hadov, korytnačiek - jedna skupina študentov; lebky krokodílov (alebo figuríny) - 1 - 2 na skupinu. Pre jedného alebo dvoch študentov sa vyžaduje:
1. Zložiteľná kostra jašterice monitora.
2. Lebka korytnačky.
3. Disekčné ihly - 2.
JOB
Zoberme si kostru jašterice monitora ako celok. Nájdite jeho hlavné časti: lebku, chrbticu, kostru párových končatín a ich opasky. Zvážte štrukturálne vlastnosti kostry hada a korytnačky. Zvážte podrobnosti o štruktúre kostry jašterice monitora a lebiek jašterice monitora, krokodíla a korytnačky. Porovnať štruktúru lebky krokodíla, monitor jašterica, had a korytnačka.
Urobte nasledujúce výkresy:
1. Lebka monitora na bočnej strane.
2. Lebka monitora jašterice zdola.
4. Predná časť jašterice monitora.
6. Zadná končatina jašterice monitora.
7. Krokodíl lebka na vrchu.
8. Krokodílna lebka zdola.
9. Lebka korytnačky na boku.
OPIS SKELETONU
Axiálna kostra. Diferenciacia axiálnej kostry alebo chrbtice na oddelenia je výraznejšia u plazov ako u obojživelníkov. Krčka chrbtice (pars cervicalis) je vždy zložená z niekoľkých stavcov, z ktorých majú dve predné časti špeciálnu štruktúru. Prvý krčný stavca sa nazýva atlas alebo atlas (atlas; obr. 81, A). Nemá telo stavcov a má tvar krúžku rozdeleného na dve časti. Na spodnej čelnej ploche tohto stavca je kĺbová dutina pohyblivo spojená s kondylom lebky (pozri nižšie). Druhá krčka stavca - epistrofia (epistrofaeus; obr. 81, B), má pred sebou veľký zubovitý proces (processus odontoideus, obr. 81, 1), čo je telo prvého krčného stavca fúzovaného s epistrofiou. Zubovitý proces voľne vstupuje do dolného otvoru atlasu. Táto štruktúra prvých krčných stavcov poskytuje väčšiu pohyblivosť hlavy. Ostatné krčné stavce majú obvyklé zariadenie (pozri nižšie); veľa z nich má krátke rebrá.
Obr. 81. stavce
A - atlas; B - epistrofia; In - hrudný stavec; G - pozdĺžny rez hrudného stavca:
1 - zubovitý proces epistrofie, 2 - stavce, 3 - horný oblúk, 4 - spinálny proces, 5 - kanál na miechu, 6 - predný artikulárny proces, 7 - zadný artikulárny proces
Hrudné a bedrové delenie nie sú celkom zreteľné a zvyčajne sa považujú za jedno delenie (pars thoracolumbalis). Hrudná oblasť je v skutočnosti časťou chrbtice, v ktorej sa rebrá siahajúce od stavcov so spodným koncom pripevňujú k hrudnej kosti. Stavce bedrovej oblasti nesú rebrá, ktoré nedosahujú hrudnú kosť. Stavce (obr. 81, 2) sú konvexné pred a konvexné zozadu; takéto stavce sa nazývajú percento. Nad stavcom sa stúpajú horné oblúky (arcus neuralis; obr. 81, 3) a končia spinálnym procesom (processusspinosus; obr. 81, 4). V kanáli tvorenom hornými oblúkmi je umiestnený miecha.
Predné procesy (processus articularis anterior; Obr. 81, 6) a zadné (processus articularis posterior; Obr. 81, 7) kĺbové procesy sa odchyľujú od prednej a zadnej časti spodnej časti horných oblúkov. Tieto spárované procesy sa spájajú s kĺbovými procesmi susedných stavcov a prispievajú k väčšej pevnosti chrbtice počas ohýbania. Na stranách tela stavcov (blízko spodku horných oblúkov) sú malé zahĺbenia, ku ktorým sa rebrá pripájajú.
Sakrálny úsek (pars sacralis) pozostáva z dvoch stavcov, ktoré sa vyznačujú silne rozvinutými priečnymi procesmi (processus transversus); sú spojené kosťami panvy. Časť chvosta (pars caudalis) je zastúpená početnými stavcami, ktoré sa postupne zmenšujú.
Táto štruktúra chrbtice je typická pre plazovú triedu, ale v niektorých skupinách prechádza sekundárnymi zmenami. Najmä v prípade hadíc, v spojitosti s redukciou párových končatín a vznikom iného typu pohybu - plazením sa po bruchu ohýbaním kmeňa - je chrbtica jasne rozdelená iba na trupovú a chvostovú časť. Všetky stavce kmeňa majú pohyblivé rebrá, ktorých spodné konce sú voľné (hrudná hadov je neprítomná) a prilieha k brušným nadržaným štrbinám.

Obr. 82. Kostra močaristej korytnačky
A - krunýř; B - plastrón:
1 - kmeň trupu chrbtice, 2 - kostrové platne, 3 - okrajové platne, 4 - kokakid, 5 - lopatka, 6 - bedrové7 - pubická kosť; 8 - ischiálna kosť
V korytnačkách sa axiálna kostra podieľa na tvorbe kostnej základne ich krunýře. Horný štít škrupiny - kapary (caparax; obr. 82, A) - sa skladá z niekoľkých radov kostných platní. Stredný (nepárový) rad týchto dosiek je vytvorený fúziou predĺžených a sploštených spinálnych a priečnych procesov stavcov kmeňa s kosťami kože; po stranách stredného radu sú párové rady kostných platní, ktoré sú spojené so zväčšenými rebrami. Okraj krunýře je tvorený kostnými doskami integumentary pôvodu. Z tohto dôvodu je chrbtová časť trupu pevne spojená s dorzálnym štítom krunýře. Krčná a kaudálna chrbtica sú pohyblivé. Súčasne sú predné krčné stavce opistocelulárne (stavce tela sú konvexné vpredu a konkávne v chrbte), zadné sú pozdĺžne a medzi týmito dvoma skupinami je jedno stavce, ktorého telo má konvexný povrch vpredu aj zozadu.
Lebka. V porovnaní s obojživelníkmi sa lebka plazov vyznačuje oveľa úplnejšou osifikáciou. Určité množstvo chrupavky sa konzervuje iba v čuchovej kapsule a vo sluchovej oblasti. Axiálne a viscerálne časti lebky sú embryá kladené osobitne, ale u dospelých zvierat predstavujú jeden celok. Zloženie lebky zahŕňa chrupavkovú (náhradnú alebo primárnu) a početnú kožu (klasickú alebo sekundárnu). Ako hlavný objekt pre štúdium je vhodné použiť lebku veľkého jašterice, monitora jašterice.

Obr. 83. Lebka monitora jašterice
A je na boku; B - zdola; B - zhora; G - späť:
1 - hlavná týlová kosť, 2 - laterálna týlová kosť, 3 - horná týlová kosť, 4 - veľká týlna foramen, 5 - týlový kondyl, 6 - predná kosť, 7 - hlavná sfenoidná kosť, 8 - otvárač, 9 - parietálna kosť, 10 - predná kosť, 11 - nosná kosť, 12 - prefrontálna kosť, 13 - preorbitálna kosť, 14 - slzná kosť, 15 - nadčasová fosília, 16 - zadná kosť, 17 - šupinovitá kosť , 18 - maxilárna kosť, 19 - maxilárna kosť, 20 - zygomatická kosť, 21 - prasknutie dolného temporálneho oblúka v dôsledku redukcie štvorhrannej zygomatickej kosti, 22 - štvorcová kosť, 23 - pterygoidná kosť, 24 - palatínová kosť, 25 - supra-pterygoidná kosť, 26 - priečna kosť, 27 - klenutá kosť, 28 - kosť zubu, 29 - uhlová kosť, 30 - kĺbová kosť, 31 - koronárna kosť
Osová lebka. V týlnej oblasti lebky sú všetky štyri týlne kosti: hlavný týl (basioccipitale, Obr. 83, 1), dva laterálne týlne (occipitale laterale; Obr. 83, 2) a horná týlna (supraoccipitale; Obr. 83, 3). Tieto kosti primárneho pôvodu obklopujú veľké týlne foramény (foramen occipitale magnum; obr. 83, 4). Dolné a bočné týlne kosti spolu tvoria jediný (na rozdiel od obojživelníkov) okcipitálny condyle (condylus occipitalis; Obr. 83, 5), artikulovaný s prvým krčným stavcom - atlasom. Spojenie hlavy s krkom pomocou iba jedného kondylu v kombinácii so štrukturálnymi znakmi prvých dvoch krčných stavcov, ktoré už boli brané do úvahy, dáva plazovej hlave významnú pohyblivosť.
V sluchovej časti chrupavkových kostí zostáva nezávislá iba párová predná kosť (prooticum; obr. 83, 6), zatiaľ čo fúzia horného ucha s hornou týlnou kosťou a zadné ucho s laterálnou týlnou kosťou.
Interorbitálne septum plazov je tenké, membránové a iba krokodíly a jašterice majú samostatné malé osifikácie, čo zjavne zodpovedá očiam klinovitých kostí. Čuchová kapsula nemá osifikáciu.
Na spodnej časti lebky, pred hlavnou týlnou kosťou, je pomerne veľká integumentárna hlavná sfénoidná kosť (fundamentenofén, obr. 83, 7). Jeho predný úzky proces je homológny s parasfenoidom (parasphenoideum), ktorý sa u plazov výrazne znižuje. V prednej časti spodnej časti lebky, pod čuchovou časťou, je párový otvárač (vomer; obr. 83, 8), tiež základného pôvodu.
Strecha lebky je reprezentovaná početnými klasickými kosťami, z ktorých niektoré padajú dole a pokrývajú lebku zo strán. Tieto zahŕňajú parietálne (parietálne; obr. 83, 9), frontálne (frontálne; obr. 83, 10) a nosné (nazálne; obr. 83, 11) kosti. Pred čelnými kosťami sú zvyčajne spárované prefrontálne (praefrontale; obr. 83, 12) a preorbitálne (praeorbitale; obr. 83, 13) kosti, ktoré sú obvykle umiestnené a pod nimi v prednej stene orbity sú spárované slzami (slzami; obr. 83, 14). ).
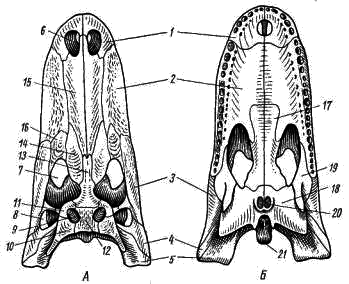
Obr. 84. Krokodília lebka (Mississippi Alligator).
A - zhora; B - spodná časť:
1 - maxilárna kosť, 2 - maxilárna kosť, 3 - zygomatická kosť, 4 - štvorcová zygomatická kosť, 5 - štvorcová kosť, 6 - vonkajšia nosná diera, 7 - obežná dráha, 8-laterálna časová fossa, 9 - vynikajúca časová fossa, 10 - šupinatá kosť, 11 - frontálna (postorbitálna) kosť, 12 - parietálna kosť, 13 - frontálna kosť, 14 - prefrontálna kosť, 15 - nosná kosť, 16 - slzná kosť, 17 - kosť palatínová, 18 - kosť pterygoidná, 19 - priečna kosť , 20 - choans (vnútorné otvory nosných dierok), 21 - týlny kondyl
Z ostatných zvyšných kostí axiálnej lebky sú osobitne zaujímavé kosti, ktoré sa podieľajú na tvorbe takzvaných časových oblúkov. Krokodíl v streche lebky mimo parietálnej kosti (obr. 84, 12) má na každej strane otvor - nadčasovú fosíliu (obr. 84, 9). Na vonkajšom okraji je horná časová fossa ohraničená zadnými frontálnymi alebo postorbitálnymi (postfrontale-postorbitale, obr. 84, 11) a šupinatými (squamosum; obr. 84, 10) kosťami. Tieto dve kosti spolu tvoria vynikajúci dočasný oblúk. Na strane lebky za obežnou dráhou sú laterálne časové jámy (Obr. 84, 8), ohraničené zvonka spodnými časovými oblúkmi. Každý dolný časový oblúk sa skladá z dvoch kostí: zygomatickej (jugale; obr. 84, 3) a štvorcovej-zygomatickej (quadrato-jugale; obr. 84, 4). Dolný časový oblúk sa spája s hornou čeľusťou: zygomatická kosť rastie do maxily a štvorcová zygomatická kosť rastie na štvorec. Tento typ lebky, ako krokodíl - s dvoma dočasnými jamami a dvoma dočasnými oblúkmi, sa nazýva diapsid (dvojramenný).
V jašterici monitora je horná časová fossa (obr. 83, 15) ohraničená úplným horným časovým oblúkom (postfrontale - squamosum; obr. 83, 16-17). V rámci dolného časového oblúka sa zmenšila štvorcová zygomatická kosť a zachovala sa iba zygomatická kosť (obr. 83). 20); bočné spánkové fosílie preto nie sú zvonku otvorené a zostávajú otvorené. Lebka monitora jašterice sa preto môže považovať za lebku typu diapsidu, ale so zníženým dolným oblúkom. U niektorých ďalších jašteríc je horný časový oblúk tiež čiastočne zredukovaný, zatiaľ čo u hadích (pozri obr. 86) sa znížia obidve dočasné oblúky (zadné predné a šupinovité kosti sa navzájom nepripájajú; obe dočasné jamy zostávajú otvorené vonku).
Hady a jašterice (šupinatý komár, Squamata) v štruktúre lebky teda patria do skupiny diapsidných (dvojramenných) plazov, ale vyznačujú sa rôznym stupňom redukcie časových oblúkov.

Obr. 85. Lebka močaristej korytnačky:
1 - falošná časná fossa, 2 - maxilárna kosť, 4 - zygomatická kosť, 5 - štvorcová zygomatická kosť, 6 - štvorcová kosť, 7 - šupinatá kosť, 8 - predná kosť, 9 - parietálna kosť, 10 - predná kosť, 11 - prefrontálna kosť, 12 - vynikajúca týlová kosť
Korytnačka nemá časové jamy a bočnú stenu strechy lebky, ktorá zvonku vymedzuje veľkú dutinu - tzv. Falošnú časovú jamu (Obr. 85, 1), ktorá sa vytvára ako výklenok v týlnej časti lebky. 85, 8), šupinatá (šupinatá; obr. 85, 7), zygomatická (jugale; obr. 85, 4) a štvorcová zygomatická (quadrato-jugale, obr. 85, 5). Tento typ lebky, zbavený pravých časných fosílií a časových oblúkov, ktoré ich obmedzujú, sa nazýva anapsidický (neživý).
Viscerálna lebka. V jašterici monitora chrupavka palatínového štvorca osifikuje, pričom v zadnej časti vytvára kvadratickú kosť (kvadratum; obr. 83, 22), na spodnom konci ktorého je pripojená spodná čeľusť; horný koniec štvorcovej kosti je pohyblivo kĺbovo spojený s axiálnou lebkou. Pred štvorcovou kosťou je pterygoid (pterygoideum; obr. 83, 23) a pred ňou je palatínová kosť (palatinum; obr. 83, 24), ktorá sa spája s maxilárnymi kosťami a vomérom. Všetky tieto kosti sú spárované; z toho iba štvorcové kosti majú pôvod v chrupavke (primárny).
Z kosti pterygoid sa vynikajúci pterygoid (epipterygoideum; obr. 83, 25) rozširuje smerom nahor. Táto párová kosť spájajúca pterygoidné a parietálne kosti je homológna s vertikálnym („vzostupným“) chrupavkovým chrupom a je charakteristická pre živé plazy jašteríc a gatterií. Okrem vynikajúcich pterygoidov sa priečne kosti (priečne; obr. 83, 26) rozprestierajú od kostí pterygoidných, ktoré sa vo svojej prednej časti spájajú s maxilárnymi kosťami. Sekundárna maxilla je reprezentovaná maxilárnymi (praemaxillare; Obr. 83, 18) a maxilárnymi (maxillare; Obr. 83, 19) kosťami. Spodná čeľusť pozostáva z primárnej kĺbovej kosti (artikula; obr. 83, 30) a základných kostí: zubu (dentale; obr. 83, 28), uhlového (angulare; obr. 83, 29), suprahlového (supraangulare; obr. 83, 27), koronárne (koronare; obr. 83, 31) a niekedy aj niekoľko ďalších malých kostí.
Na premaxilárnych, čeľustných a zubných kostiach plazov (okrem korytnačiek) sa nachádzajú jednoduché kužeľovité, niekedy mierne ohnuté chrbtové zuby, ktoré dorastajú až po okraj zodpovedajúcej kosti.
Podjazykový oblúk, podobne ako obojživelníci, úplne stratil funkciu zavesenia. Horný prvok hyoidného oblúka (hyomandibulárny) je súčasťou stredného ucha vo forme tyčinkovitej sluchovej kukly - stonky (stapes seu columella) a zvyšok z nich spolu so zvyškami predných žiabrových oblúkov tvorí hyoidný aparát.
Opísaná štruktúra viscerálnej lebky je všeobecne typická pre všetky plazy. V niektorých skupinách však existujú odchýlky od tejto schémy, ktoré sa týkajú najmä špecifík biologie týchto skupín.

Obr. 86 Lebka jedovatého hada:
1 - maxilárna kosť, 2 - maxilárna kosť. 3 - palatínová kosť, 4 - pterygoidná kosť, 5 - priečna kosť, 6 - štvorcová kosť, 7 - šupinatá kosť, 8 - zadná kosť, 9 - jedovatý zub, 10 - predná kosť, 11 - nosná kosť, 12 - zubná kosť. 13 - uhlová kosť, 14 - kĺbová kosť
U hadí sú nielen pohyblivé štvorcové (obr. 86, 6), ale aj šupinaté (obr. 86, 7), ako aj pterygoidové (obr. 86, 4) a palatínové (obr. 86, 3) kosti. Posledné dva majú ostré zuby. Priečne kosti (obr. 86, 5) v hadoch slúžia ako páky, ako to bolo, prenášajúce pohyb pterygoidných kostí na maxilárne kosti (obr. 86, 2), ktoré sú zase veľmi pohyblivé. Celý tento systém pohyblivých kĺbov prispieva nielen k extrémne širokému otvoru v ústach, ale tiež zaisťuje nezávislé pohyby pravej a ľavej polovice čeľusťového aparátu, keď sa korisť tlačí do hrdla striedavým zachytením. To umožňuje hadom prehltnúť pomerne veľkú korisť (presahujúcu hrúbku tela hada). Jedovaté hady na maxilárnych kostiach majú pohyblivo pripojené ostré, dozadu zakrivené jedovaté zuby (obr. 86, 9), ktoré majú na prednom povrchu vnútorný kanálik alebo drážku, cez ktorú prúdi jed z jedovatých žliaz umiestnených na spodnej časti zubu, keď je uhryznutý.
Lebka krokodíla (obr. 84) sa vyznačuje tým, že zuby nerastú až na okraj zubných, maxilárnych a maxilárnych kostí, ako u iných plazov, ale sú umiestnené v špeciálnych dutinách (dierach alebo alveolách) týchto kostí - zuboch tecodont. Ďalším znakom viscerálnej lebky krokodílov je sekundárny tvrdý patro, ktorý oddeľuje ústnu dutinu od nosohltanu.
Palatínové procesy predaxilárnych (obr. 84,7) a maxilárnych (obr. 84, 2) kostí, ako aj palatínových (obr. 84, 17) a kostí pterygoidových (obr. 84, 18) sa podieľajú na tvorbe sekundárneho tvrdého podnebia. V dôsledku tvorby tvrdého podnebia sa sekundárne výbery (obr. 84, 20) prenášajú späť a nachádzajú sa v kostiach pterygoidu nad hrtanom. Tvorba sekundárneho tvrdého podnebia súvisí s povahou krokodílového životného štýlu: priamy kontakt hrtanu so choanom otvára možnosť nepretržitého dýchania pri jedle a keď krokodíly spočívajú v plytkej vode, pričom sa z úst vychádzajú zvýšené nosné dierky, zatiaľ čo ústna dutina je naplnená vodou.
Párové končatiny a ich opasky. Ramenné pásy plazy sa skladajú z typických kostí: lopatka umiestnená viac dorsálne (lopatka; obr. 87, 1) a korakoid (coracoideum; obr. 87, 3) smerujúce k ventrálnej strane. Obidve tieto kosti sa podieľajú na tvorbe kĺbovej dutiny (obr. 87, 4) na pripevnenie prednej končatiny. Dorsálna k lopatke je široká sploštená supraskapulárna chrupavka (chrupavka suprascapularis; obr. 87, 2) a pred kortexom je chrupavková prokokozida (chrupavka procoracoidea; obr. 87, 5). Existuje dobre vyvinutá sternum (sternum, obr. 87, 6), ku ktorej je pripojených niekoľko rebier (kosta, obr. 87, 7). Na rozdiel od obojživelníkov tak plaz vytvára hrudník a ramenný opasok má podporu v axiálnom skelete. Na prednej strane hrudnej kosti je integumentárna kosť v tvare písmena T - suprasternum (episternum; obr. 87, 8), pred ňou - tiež integumentary kosti - klavicle (clavicula; obr. 87, 9). Vonkajšie konce kľúčnej kosti sú pripevnené k lopatkám lopatiek a vnútorné sú spojené s vetvami hrudníka. Kĺčiky a suprasternum (chýbajú u obojživelníkov) zvyšujú pevnosť spojenia pravej a ľavej časti ramenného pletenca.

Obr. 87. Ramenný opasok jašterice monitora (pohľad zdola):
1 - lopatka, 2 - supraskapulárna chrupavka, 3 - korakoidná, 4 - kĺbová dutina na hlave ramena, 5 - proorakálna chrupavka, 6 - hrudná kosť, 7 - rebrá, 8 - suprasternum, 9 - kľúčná kosť
U hadov je ramenný opasok úplne zredukovaný a v korytnačkách sú kľúčové kosti a suprasternum zahrnuté v kostiach brušného štítu (plastrón; pozri obr. 82, B) krunýča, pričom medzi nimi tvoria klinové platne prednej a nepárovej kosti.

Obr. 88. Panvový opasok jašterice monitora (pohľad zdola):
1 - ilium, 2 - ochlpenie kosti, 3 - sedacie kosti, 4 - acetabulum (kĺbová fossa) pre hlavicu stehennej kosti, 5 - sakrálne stavce
Panvový pás sa skladá z dvoch symetrických polovíc spojených pozdĺž stredovej čiary chrupavky. Každá polovica je zložená z troch kostí; umiestnené na chrbte iliak (ilium; obr. 88, 1), umiestnené na ventrálnej strane ochlpenia (pubis; obr. 88, 2) a sedacieho nervu (ischium, obr. 88, 3). Všetky tieto kosti sa podieľajú na tvorbe kĺbovej fosílie (acetabulum; obr. 88, 4), ku ktorej je pripojená zadná končatina. Panva plazov je uzavretá: pravá a ľavá ochlpenie na krku a ischiálne kosti na boku sú navzájom spojené.

Obr. 89. Konce jašterice monitora
A - predné; B - späť:
1 - humerus, 2 - ulna, 3 - polomer, 4 - zápästie, 5 - metakarpus, 6 - falanga prstov, 7 - medzikarpálny kĺb, 8 - stehenná kosť, 9 - holenná kosť, 10 - holenná kosť, 11 - patella, 12 - tarzus, 13 - medzistranný kĺb, 14 - metatarz
Končatiny plazov sa stavajú podľa typického vzoru končatín suchozemských stavovcov. Proximálna časť prednej končatiny (Obr. 89, A) je predstavovaná jednou kostou - humerus (humerus; Obr. 89, 1), nasledovanou predlaktím, ktoré pozostáva z dvoch kostí - ulnár (ulna; Obr. 89, 2) a polomer (polomer); Obr. 89, 3). Zápästie (carpus, obr. 89, 4) pozostáva z relatívne malých kostí, ktoré sa zvyčajne nachádzajú v dvoch radoch; na ich strane je ďalšia hruškovitá kosť, odobratá po zvyšok šiesteho prsta. Metacarpus (metacarpus; obr. 89, 5) sa skladá z piatich podlhovastých kostí, na ktoré sú pripevnené falangy piatich prstov (dighala falanges; obr. 89, 6).
Posledné falangy majú pazúry. Kĺb, zabezpečujúci pohyb ruky, u plazov neprechádza medzi kosťami predlaktia a proximálnym radom kostí zápästia (ako u obojživelníkov), ale medzi proximálnymi a distálnymi radmi kostí zápästia. Takýto kĺb sa nazýva medzikarpálny (obr. 89, 7).
V zadnej končatine je kĺbový člen - stehenná kosť (femur; obr. 89, 8) kolenný kĺb s holennou kosťou pozostávajúcou z dvoch kostí holennej kosti - veľkej (holennej kosti; Obr. 89, 9) a malej (holennej kosti; Obr. 89, 10). Nad predným povrchom tohto kĺbu je malá kosť - patella (patella; Obr. 89, 11). V tarzuse (tarzus; obr. 89, 12) sa proximálna rada kŕstkov spája alebo takmer bez pohybu sa spája s kosťami dolnej končatiny a kosťice distálnej rady sú tiež úzko spojené a čiastočne fúzované s metatarzálnymi kosťami. Z tohto dôvodu je kĺbová plocha tu umiestnená nie medzi dolnou časťou nôh a chodidla, ale medzi proximálnymi a distálnymi radmi tarzálnych kostí. Takýto kĺb je charakteristický pre plazy a nazýva sa medzistupňový kĺb (Obr. 89, 13). 89). Metatarsus (metatarsus; Obr. 89, 14) pozostáva z piatich podlhovastých kostí, na ktoré sú pripevnené falangy piatich prstov (falanges digitorum; Obr. 89, 6). Koncové falangy nesú pazúry.
ZÁVER
Plazy (Reptilia) - prvá trieda skupiny pravých suchozemských stavovcov (Amniota), ktorej predstavitelia majú komplex úprav, aby mohli žiť na zemi bez akéhokoľvek spojenia s vodným prostredím.
Vyššie bolo povedané, že úzke spojenie predkov plazov - obojživelníkov - s vodnými útvarmi (alebo aspoň s miestami s vysokou vlhkosťou) bolo spôsobené predovšetkým dvoma biologickými črtami: vodnou reprodukciou a dualitou dýchacieho systému (pľúca a koža). Tieto vlastnosti sa odrážajú na povahe metabolizmu vody: obojživelníci, ktorí žijú vo vlhkom prostredí, nemajú morfofyziologické mechanizmy na zníženie straty telesnej vlhkosti. Úplný rozvoj pôdy je nemysliteľný bez zásadných premien, ako sú rozmnožovanie, dýchanie a metabolizmus vody.
Realizácia týchto transformácií charakterizuje triedu plazov.
Najšpecifickejšie črty tejto triedy (a iné amnioty) ako primárne suchozemské stavovce sú spojené s prispôsobením sa životu v relatívne suchom atmosférickom vzduchu. Špecifické štrukturálne vlastnosti plazov sú spojené predovšetkým s prispôsobením sa hospodárnejšiemu použitiu vlhkosti. Koža plazov je suchá a bez žliaz (s výnimkou malého počtu špecializovaných žliaz, ktoré vylučujú relatívne silné tajomstvo). Vonkajšie vrstvy epidermy sú zastúpené odumretými bunkami naplnenými nadržanou hmotou (keratogialín).
Táto stratum corneum chráni telo pred vyschnutím a tiež výrazne zlepšuje ochranu pred chemickým a mechanickým poškodením. Pod stratum corneum je malpigiánna vrstva epidermy zastúpená živými bunkami. Keď sú odumreté bunky opotrebované, sú nahradené bunkami zo spodných vrstiev, v protoplazme, z ktorej sa najskôr objavia oddelené zrná keratogialínu, ktoré následne vyplnia celú bunku. Okrem všeobecnej keratinizácie hornej vrstvy epidermy sa plazy vyznačujú tým, že v koži sa tvoria rôzne formy rohovitých šupín, ktoré pokrývajú povrch kože. Váhy poskytujú mechanickú ochranu pokožky a prispievajú k ešte väčšiemu zníženiu straty vlhkosti v tele. niektoré moderné plazy (napríklad krokodíly) majú kostnaté platničky pod nadržanými váhami.
Všetky tieto transformácie, biologicky prospešné z hľadiska udržiavania ekonomickejšej vodnej rovnováhy tela, boli možné predovšetkým preto, že plazová koža sa nepodieľa na dýchaní, a to kvôli postupnému vývoju dýchacieho systému (pozri nižšie).
Pri vývoji vyšších suchozemských stavovcov má veľký význam transformácia vylučovacieho systému. Na rozdiel od predtým študovaných tried majú plazy (podobne ako iné Amnioty) panvové obličky (metanefy). Na rozdiel od obličiek kmeňa (mezonephros) anamnéza vylučovacích kanálikov panvovej obličky úplne stratila kontakt s telesnou dutinou; metabolické produkty vstupujú do vylučovacích kanálikov iba z plexu krvných kapilár - Malpighian glomerulus. Je uzavretá v dutom výhonku konca trubice - Bowmanovej kapsule. To vedie k tomu, že v procese izolácie produktov disimulácie telo stráca menej vody (tekutina v dutine sa nevylučuje). Krvné cievy vychádzajúce z malpighiánskeho glomerulu tvoria druhú sieť kapilár okolo steny vylučovacej trubičky a potom prúdia do väčších ciev žilového systému. Prostredníctvom tejto druhej siete kapilár sa voda, uhľohydráty a niektoré soli re-absorbujú do krvi, takže sa uvoľňuje tzv. Sekundárna moč, ktorá je koncentrovanejšia a má iné zloženie ako „primárna moč“ tvorená v bowmanových kapsulách.
Panvová (metanefrická) oblička má svoj vlastný močovod, ktorý nie je homológny s Müllerovými alebo Wolfovými kanálmi. Vlk je zachovaný iba u mužov, kde je cez epididymis spojený s pohlavnou žľazou a hrá úlohu vas deferens. Muellerov kanál je znížený. Ženy naopak zachovávajú iba Müllerove kanály; vykonávajú funkciu vajcovodov a otvárajú sa do telovej dutiny s otvormi - lievikmi vajcovodov. Vlkové kanály u žien sú znížené.
Hlavným znakom vylučovacieho systému vyšších suchozemských stavovcov je teda strata komunikácie medzi obličkami a telesnou dutinou a posilnenie systému spätnej absorpcie vody z primárneho moču do krvi. Tieto vlastnosti zabezpečujú vykonávanie vylučovacej funkcie s minimálnou spotrebou vody, ktorá má pre obyvateľov krajiny, ako už bolo uvedené, veľký biologický význam. K tomu môžeme dodať, že medzi suchozemskými stavovcami je intenzita všeobecnej filtrácie a spätnej absorpcie vody u zvierat žijúcich v rôznych podmienkach rôzna. Takže medzi plazmi, krokodíly a korytnačky (väčšinou sladkovodné formy) majú dobre vyvinuté glomeruly a vylučujú tekutý moč, zatiaľ čo v jaštericiach a hadoch sú glomeruly výrazne slabšie a moč je tuhý alebo polotekutý.
Celý komplex uvažovaných prvkov, ktorý poskytuje hospodárnejšiu vodnú bilanciu plazov, spočíva na konečnom oddelení vyšších suchozemských stavovcov od vodného prostredia a ich rozšírenej populácie rôznych pôdnych biotopov.
Spolu s týmito znakmi sa plazy vyznačujú celým radom transformácií, ktoré majú pri vývoji stavovcov spoločnú progresívnu hodnotu.
Dýchací systém sa výrazne zmenil. Objavila sa priedušnica, ktorej dĺžka závisí od dĺžky krku. Pľúca plazov sa vyznačujú zložitejšou vnútornou štruktúrou ako obojživelníky: majú vnútorné priečky, ktoré ich delia na oddelenia. Z týchto priečok sa priečky zase odchyľujú, vyvíjajú sa do rôznych stupňov v rôznych skupinách. Medzi septou sa tvoria intrapulmonárne dýchacie cesty, ktoré slúžia ako pokračovanie priedušiek av niektorých prípadoch sa delia na intrapulmonálne priedušieky druhého rádu. Vnútorné steny komôr tvorené celým týmto systémom priečnych nosníkov majú bunkovú štruktúru, takže v dôsledku toho pľúca spravidla získajú komplexnú špongiovú štruktúru. Takéto zariadenie pľúc významne zvyšuje celkový povrch dýchacích ciest, a teda aj intenzitu dýchania. Vďaka týmto zariadeniam je potreba kyslíka plazovho organizmu úplne zabezpečená samotným pľúcnym dýchaním.
Je mimoriadne dôležité, aby sa akt dýchania plazov vykonával aktívnym rozširovaním a sťahovaním brušnej dutiny pôsobením brušných a brušných svalov. Tento typ dýchania poskytuje intenzívnejšie vetranie pľúc veľkým objemom vzduchu; jeho vzhľad je spojený s vývojom pohyblivej hrudníka.
V obehovom systéme plazov sa vyskytujú významné progresívne transformácie (v opise obehového systému bola uvedená schéma na oddelenie venóznych a arteriálnych krvných tokov v srdci plazov).
V dôsledku výskytu neúplného septa v komore a nezávislého odchodu z rôznych oddelení hlavných kmeňov tepien (čo je spojené so znížením arteriálneho kužeľa) sa dosiahne pomerne úplné oddelenie arteriálnych a venóznych prietokov krvi. V čase komorovej systoly sú jej vnútorné dutiny nejaký čas izolované; zatiaľ čo venózna krv vstupuje do pľúcnej tepny, arteriálna krv vstupuje do pravého oblúka aorty a arteriálna venózna prímes vstupuje do ľavého oblúka aorty. Pretože karotická aj subklaviálna artéria pochádzajú z pravého oblúka aorty, hlave a prednej časti tela sa dodáva takmer čistá arteriálna krv (napríklad v korytnačkách v krčných tepnách saturácia kyslíkom v krvi prekročila 90% v polovici prípadov). Plavidlá siahajúce pod spojenie aortálnych oblúkov prenášajú krv obsahujúcu určitú žilovú nečistotu do orgánov zadnej polovice tela.
Horizontálne septa, ktoré delí komoru srdca plazov na dve hlavné komory, nie je homologická s interventrikulárnym septom vtákov a cicavcov. Zo všetkých skupín plazov iba v krokodíloch sa v komore objavuje nové - zvislé - septum, ktoré nahrádza starú horizontálnu. Táto vertikálna septa úplne rozdeľuje srdcovú komoru krokodílov na dve izolované komory: pravú (venóznu) a ľavú (arteriálnu). Srdce krokodílov teda možno nazvať štvorkomorové: má dve predsiene izolované od seba a dve úplne oddelené komory. Spolu s úplným oddelením komôr si však krokodíly zachovávajú obidve aortálne oblúky, ktoré tvoria krížovú charakteristiku všetkých plazov: pravá strana začína vľavo a ľavá v pravej komore. Na križovatke je tzv. Panniciusový otvor spájajúci dutiny oboch aortálnych oblúkov.
Karotická a subklaviálna artéria, rovnako ako všetky plazy, sa tiahne od pravého oblúka aorty. Ľavý aortálny oblúk obiehajúci okolo srdca dodáva krvu do tenkého čreva a cez tenký kanálik sa pripája k pravému oblúku aorty, čím vzniká miecha. Podľa niektorých správ sú na spodnej časti ľavého aortálneho oblúka chlopne a systém chrupavky, ktorý zabraňuje vstupu žilovej krvi do nej pomocou komorovej systoly.
Niektorí autori vysvetľujú zachovanie aortálnych oblúkov a ich spojenie v mieste priesečníka s ekologickými charakteristikami krokodílov: zatiaľ čo pod vodou cirkuluje pľúcny obeh trochu krvi, pretože dýchanie je pozastavené a prebytok krvi z pľúcneho obehu prechádza do veľkého kruhu, čím podporuje jeho krvné cievy normálny krvný tlak. Je tiež možné, že zachovanie botallalkov (cievy spájajúce aortálne oblúky s pľúcnymi tepnami) v korytnačkách vedúcich k vodnému životnému štýlu je rovnako dôležité. Rovnako ako karotidové kanáliky (spojenie aortálnych oblúkov s karotickými tepnami) hadíc a jašteríc, aj tieto útvary sú základmi arteriálnych oblúkov vzdialených predkov plazov, ktoré sa znížili, keď sa zmenilo dýchanie žiabrov. Najprimitívnejšie zo živých plazov, klobúky, sú krčné a duktálne kanály.
centrálnej nervový systém plazy sa vyznačujú zvýšením relatívnej veľkosti mozgu a progresívnym vývojom predných mozgových hemisfér, na streche ktorých sa najskôr objavuje povrchová sivá hmota - mozgová kôra (stále slabo vyvinutá).
Medzi progresívnymi znakmi kostry plazov je dôležitý vzhľad uzavretej hrudníka. Okrem už naznačenej hodnoty tejto transformácie pri zintenzívnení dýchania hrudník zabezpečuje spojenie ramenného pletenca s axiálnym skeletom, a tým jeho pevnejšie fixovanie, čo vedie k zvýšeniu podporná funkcia končatiny. U hadov, ktorí už druhýkrát prišli o párové končatiny, sa zníži aj ich opasok.
Vďaka jedinému týlnemu kondylu sa špecifická štruktúra prvých krčných stavcov, ktorá umožňuje atlasu otáčať sa okolo procesu podobného zubu epistrofie, a objavenie sa mnohých krčných stavcov, ktoré sú vzájomne pohyblivo kĺbovo spojené, výrazne zvýšila mobilita hlavy plazov. To má veľký biologický význam pri orientácii, vyhľadávaní a získavaní potravy atď.
Evolučné zmeny na streche lebky sú veľmi významné. U najstarších plazov bola horná a bočná stena lebky zložená z početných klasických kostí a mala vzhľad pevného krunýře. Taká lebka sa volala prešívaná. Ďalší vývoj prebiehal cestou postupného zmenšovania integumentárnych kostí prešívanej lebky takým spôsobom, že namiesto redukovaných kostí sa vytvorili depresie (dočasné jamy), ohraničené relatívne tenkými kostnými mostíkmi (dočasné oblúky). Tieto premeny lebky na jednej strane viedli k jej úľave, čo je dôležité, keď žijete v oveľa menej hustejšom prostredí ako vo vodnom ovzduší; na druhej strane boli použité priehlbiny vytvorené v mieste zmenšených kostí strechy lebky. Prispôsobiť svaly čeľustí. Vývoj svalov čeľuste priamo súvisí so zvýšením výživovej aktivity.
Vo väčšine skupín plazov sa počas redukcie sekundárnej strechy lebky vytvorili dve časové jamy: horná, ohraničená zvonka horným časovým oblúkom (zadné predné a šupinaté kosti) a bočná ohraničená nad hornou a vonkajšou dolnou (zygomatické a štvorcové zygomatické kosti). Plazy s týmto typom lebky patria do skupiny diapsidov (bipedálne). U niektorých predstaviteľov tejto skupiny viedla redukcia kostí ďalej v smere straty jednej alebo dokonca obidvoch časných oblúkov. Typicky výrazná diapsidná lebka je charakteristická pre živé tvary pre klobúky a krokodíly. U mnohých jašteríc je dolný časový oblúk čiastočne zredukovaný av niektorých horný. Obidve oblúky sú u hadov redukované. Vtáky sa vyvinuli z plazov plazov (u týchto zvierat sa úplne zachoval spodný časový oblúk, horný sa však zmenšil).
U niektorých plazov sa vývoj časovej oblasti lebky pohyboval po ceste formovania iba jedného časného výbežku, obmedzeného na jediný dočasný oblúk zložený z zygomatických a šupinatých kostí. Pretože zygomatická kosť je charakteristickým prvkom dolnej a šupinatej kosti je horný časový oblúk diapsidnej lebky, takýto oblúk sa nazýva zmiešaný a plazy s takou štruktúrou lebky patria do skupiny synapsidov (plazy so zmiešaným oblúkom). Medzi modernými plazmi neexistujú žiadne synapsidné formy. Tento typ lebky je charakteristický pre potomkov plazov synapsidných - cicavcov.
V jednej z vetiev primitívnych plazov išlo o zmenšenie vnútorných kostí strechy lebky inak: tvorba zárezu v týlnej časti spodnej časti lebky viedla k vytvoreniu na každej strane jednej dutiny - falošnej časovej fosílii, ktorá bola na vonkajšej strane pokrytá širokým kostným mostíkom z predných, šupinatých a štvorcových kostí. Takéto plazy patria do skupiny anapsidov (neprimeraných). Medzi živými plazmi patria korytnačky do tejto skupiny.
Súbor progresívnych rysov plazov viedol k všeobecnému zvýšeniu energie životných aktivít týchto zvierat a objaveniu sa medzi nimi rôznych foriem života, odrážajúcich prispôsobenie rôznym životným podmienkam. Na rozdiel od obojživelníkov, ktoré jedia výlučne krmivo pre zvieratá, existujú medzi plazmi bylinožravé formy (napríklad suchozemské korytnačky). Avšak najmodernejšie plazy sú mäsožravce. Väčšia mobilita v porovnaní s obojživelníkmi im umožňuje loviť mobilnú korisť, niekedy dosahujú významnú veľkosť. V mnohých prípadoch úspech lovu uľahčuje množstvo špeciálnych zariadení: jedovaté zuby u niektorých druhov hadov, ochranná farebná charakteristika mnohých plazov atď.
Väčšina plazov sú suchozemské zvieratá. Na zemi sa vyskytujú rôzne druhy plazov v najrôznejších podmienkach: v púšti, húštiach močaristej vegetácie, lesoch, medzi skalami atď. Všetky tieto druhy majú vhodné zariadenia na pohyb (prsty ohraničené rohovými platničkami v pieskových púšťach alebo štetinách rohov, ktoré umožňujú lezenie) zostaňte na strmých povrchoch), aby ste predišli útokom predátorov (hrabanie v piesku, autotómia chvosta atď.) atď.
Mnoho plazov opäť prešlo na vodný život (morské a sladkovodné korytnačky, krokodíly, vodné hady). Niektoré z týchto druhov trávia takmer celý svoj život vo vode, ale kvôli reprodukcii nevyhnutne chodia na pôdu. Morské hady si vyvinuli schopnosť oviparous, čo im umožnilo opäť sa úplne prepnúť na vodný spôsob života (medzi vyhynutými plazmi, ichthyosaury, ktoré tiež majú schopnosť oviparous), viedli úplne vodný spôsob života).
Plazy dosiahli najväčšiu rozmanitosť foriem života počas evolučného rozkvetu tejto triedy - v mezozoickej ére. Z fosílnych plazov sú známe bylinožravce a mäsožravce (rôzne skupiny dinosaurov), hlboko špecializovaní obyvatelia vodného prostredia (ichthyosaury) a lietajúce formy (pterodaktyly, ramphorinchs), ako aj mnohé ďalšie biologické skupiny. Všeobecné ochladzovanie klímy, ktoré nastalo po druhohorách, neprispelo k ďalšej prosperite plazov: rovnako ako obojživelníky sú plazy poikilotermálne a počas chladného úderu nemôžu dlho držať vysokú teplotu tela.
Väčšina druhov moderných plazov žije v horúcom podnebí; druhy obyčajné v miernom pásme trávia významnú časť roka v omračovaní. Za týchto podmienok sa ukázali jasné výhody na strane ich „potomkov“ - vtákov a cicavcov, pri ktorých vývoji sa objavila nová hodnotná kvalita - homeoterma. Schopnosť udržiavať aktívny život v širokom rozmedzí teplôt okolia je jedným z hlavných dôvodov určovania prevládajúcej polohy vtákov a cicavcov v modernej faune stavovcov.
Ďalšie čítanie
Gurtovoi N. N., Matveev B. S. Dzerzhinsky F. Ya. Praktická zootómia stavovcov. Obojživelníci, plazy. M., 1978
Terentyev P. V. Herpetology. M., 1961.
Schmalhausen I. I. Základy porovnávacej anatómie stavovcov. M., 1947
Chrbtica kombinuje 4 oddelenia: krčné, bedrové, hrudné, sakrálne a kaudálne. Stavce tela sú konvexné vpredu a konvexné v chrbte - celé stavce.
Horné oblúky končia dobre definovaným odstredivým procesom; pár krátkych predných kĺbových procesov sa odchyľuje od predného okraja nadradeného oblúka a pár zadných kĺbových procesov odchádza od zadného okraja toho istého oblúka. U dospelých nie sú žiadne známky zvyškov akordu.
Kostra jašterice (podľa Bram):
1 - kľúčna kosť, 2 - lopatka, 3 - plece, 4 - polomer, 5 - ulna, 6 - zápästie, 7 - zápästie, 8 - falanga prstov, 9 - rebrá, 10 - panva, 11 - stehno, 12 - veľké holenná kosť, 13 - holenná kosť, 14 - tarzus, 15 - metatarz, 16 - kaudálne stavce
krčnej chrbtice zastúpené ôsmimi stavcami. Predná strana, nazývaná atlas alebo atlas, má kruhový tvar charakteristický pre všetky amnioty. Na prednej strane jeho spodnej časti je nepárový kĺbový povrch, pomocou ktorého sa stavca kĺbovo lebky. Atlasová diera je rozdelená zväzkom na dve polovice - hornú a dolnú. Miecha prechádza cez hornú dieru a dentoidný proces (processus odontoideus) vstupuje do dolnej diery, ktorá je pevne spojená, ale nie je spojená s druhým stavcom, ktorý sa nazýva epistrofa (epistrofa). Ako dokazujú embryologické údaje, dentát je oddeleným telom prvého stavca. Atlas je teda mobilný vzhľadom na svoje vlastné telo. Posledné tri krčné stavce nesú pár dobre vyvinutých krčných rebier, ktoré sú pohyblivo pripevnené k stavovcom, postupne stúpajú dozadu a voľne končia bez dosiahnutia hrudnej kosti. Krčková časť plazov sa všeobecne líši od krčných obojživelníkov nielen vo veľkom počte stavcov, ale aj v mobilnejších: artikulácia prvého stavca s lebkou a druhým stavcom. Výsledkom je, že hlava dostáva oveľa väčšiu slobodu pohybu.
Bedrovej a hrudnej chrbtice jašterica obsahuje 22 stavcov. Všetky sú vybavené pohyblivými rebrami, ktorých dĺžka sa smerom dozadu postupne znižuje. Každé rebro sa skladá z hornej kosti a spodnej chrupavky. Prvých päť bedrových hrudných stavcov (v skutočnosti hrudných stavcov) má zvlášť dlhé rebrá, ktoré sa pripájajú k hrudnej kosti a tvoria hrudník (Thorax). Sternum (sternum) je chrupavka a má tvar kosoštvorcovej platne. Embryonálny vývoj amniotskej hrudnej kosti ukazuje, že je tvorená prerezaním distálnej častikoniec rebier, a preto nie je homológny s „hrudnou časťou“ obojživelníkov, vytvorenou nezávisle. V jašterici je už načrtnuté rozdelenie trupu chrbtice na hrudnú a bedrovú chrbticu, ktoré je pre amniotov mimoriadne charakteristické.
Chvostová časť pozostáva z početných stavcov (niekoľko desiatok). Predné sú podobné tým sakrálnym, ale ich telá sú dlhšie, priečne procesy sú tenšie a odstredivé procesy sú dlhšie. Vzadu dozadu sa kaudálne stavce postupne zmenšujú, strácajú svoje procesy a menia sa na malé tyčinkovité kosti.
Viac zaujímavých článkov
Vývoj a komplikácia motorického systému plazov je spôsobená prechodom na dominantné biotopy na súši a potrebou prekonať nerovnosti jeho reliéfu. Hlavné zmeny v podpornom aparáte plazov v porovnaní s obojživelníkmi sú spojené zvýšená mobilitapotrebné pre úspešné hľadanie potravy, rozmnožovania, ochrany pred nebezpečenstvom atď. Odráža sa to stvrdnutie lebky, väčšia diferenciácia axiálnej kostry, posilnenie končatín a ich pásov, ďalší rozvoj špecializovaných svalov.
Kostru plazov charakterizuje významný vývoj kostí prvky a súvisiace redukcia chrupavky útvary. Strecha, boky a spodok mozgovej lebky sa skladajú z obzvlášť veľkého počtu klasických osifikácií. Významné zmeny a viscerálne oddelenie. Čeľusťový prístroj sa stal masívnejším, kosti hornej čeľuste sú hlboko zakorenené horná časť oddelenie mozgu. Zároveň lebka získala mobilitu v spojení mozgovej a viscerálnej sekcie. Takže hady spodná časť lebka je komplex pákový kĺbový systém, čo vám umožňuje prehltnúť celú veľkú korisť (boa, vaječné hady, atď.). Prispejte k tomu časové jámy a oblúky, byť miestom pripútania silných a elastické svaly, Kostra hlavy sa kĺbovo spája s chrbticou jeden kondyltvorené kosťami šije. Lebka sa spravidla stala silnejšou a zároveň ľahšou.
Komplikácia diferenciácie chrbtice sa prejavuje zvýšením počtu stavcov v oddelení kmeňa. Mimoriadny význam zväčšenie krčka maternice Oddelenie, ktoré určuje pohyblivosť hlavy, čo zase zvyšuje orientáciu na zemi a pomáha zrýchľovať pohyb. Spojením prednej časti rebier s hrudnou kosťou sa tvorí hrudník, Vyvíja sa v oblasti hrudníka medzirebrové svaly, ktorý určuje dýchací mechanizmus u mnohých druhov.
končatina väčšina plazov je krátka, pripútaná k pásom končatín a majú špecializované svaly. V oblasti spojenia predlaktia s kefou sa vytvára medzikarpálny kĺb, Podobná formácia nastáva v zadných končatinách, kde je vytvorená časť členku intertarsálna artikulácia, Tieto útvary významne prispievajú k posilneniu a mobilite dolných končatín. Koncový pás pri zachovaní významného množstva chrupavky sú končatiny spojené axiálna kostra.
Plazy, ktoré stratili končatiny, ich opasky a hrudník (hady, beznohé jašterice), majú dlhé rebrá, silné trupové a podkožné svaly. U mnohých plazov (korytnačiek) je strata rýchlosti pohybu spojená s vývojom špeciálneho ochranného zariadenia - pevná škrupinahnusný pohyb. Štruktúrne vlastnosti kostry sú tiež v sekundárne vodné druhy (morské korytnačky, morské hady, leguány, krokodíly).
Po oboznámení sa so základnými charakteristikami kostry môžeme pristúpiť k podrobnejšiemu skúmaniu štruktúry jej jednotlivých častí.
lebka
Lebka je predĺžená, s významným obsahom kostných prvkov (Obr. 51). Určuje sa konkrétny tvar lebky, pomer veľkostí jej jednotlivých častí v rôznych plazoch druh energie a metóda snímania potravy.
Mozgová lebka stvrdnuté zvýšením počtu kostí a ich pevného spojenia so sebou.
Strecha lebky tvorené pármi nosné, predné predné, predné, zadné predné (postorbitálne) a parietálne kosti. bezkonkurenčná interparietal kosť má parietálny foramen autorita. Po stranách týchto prvkov rastú slzné a infraorbitálne kostiktoré rámujú hornú časť obežnej dráhy očí.
dno lebky hlavná sfenoidná kosť s východiskovým parasphenoid a dvojité otváračepo stranách ktorých leží choanae. Prvky viscerálnej časti sa tiež podieľajú na tvorbe spodnej časti mozgovej lebky. Kosti pterygoidov spojiť s štvorec, ktoré sú hady a jašterice pohyblivo spojené s mozgovou lebkou a pôsobia ako zavesenie čeľusťového aparátu. medzi pterygoids a čeľustnejsa nachádza priečna kosť, čo ďalej posilňuje lebku.
okcipitální pozostáva zo štyroch kostí - hlavné, supraoccipitálne a párové bočné. V dôsledku ich vzájomného rastu jeden týlny kondyl, V dolnej časti oddelenia je týlny foramen na spojenie mozgu a miechy.
Bočná časť lebky tvoria tri páry ušné kosti (anteroposterior, superior auricular a posteric), ktorí sú zapojení do vzdelávania časové doly.Predné a zadné kosti sú spojené s kosťami v týlnej oblasti. Vedľa nich sú temporálnej, mierkahorné časti alar kostí. spárované stĺpovité kosti pripojte strechu lebky k spodnej časti. Veľké očné jamky tvoria susedné kostné prvky - infraorbitálny, postorbitálny, zygomatický, slzný. Interorbitálne septum tvorené výrastkami frontálnych a parietálnych kostí. Na prednú časť mozgovej lebky maxilárne a intermaxilárne kosti sú vysoké. čuchovýoblasť lebky chrupavka.
Obr. 51. Lebka jašterice nad (A), pod (B), stranou (C) a jej spodnou čeľusťou:
1 - týlny foramen, 2 - laterálna týlna kosť, 3 - horná týlna kosť, 4 - hlavná týlna kosť, 5 - týlny kondyl, 6 - hlavná sfenoidná kosť, 7 - vomér, 8 - choans, 9 - vestigiálny parasfenoid, 10 - parietálny kosť, 11 - otvor pre parietálne oko, 12 - predná kosť, 13 - nazálna kosť, 14 - maxilárna kosť, 15 - maxillary kosť, 16 - prefrontálna kosť, 17 - slzná kosť, 18 - infraorbitálna kosť, 19 - orbitálna kosť, 20 - šupinatá kosť, 21 - časná kosť, 22 - zygomatická kosť, 23 - nosná diera, 24 - štvorcová kosť, 25 - palatínová kosť, 26 - pterygoidná kosť, 27 - stĺpcová kosť, 28 - priečna kosť, 29 - kĺbová kosť, 30 - zubná kosť, 31 - uhlová kosť, 32 - klenutá kosť, 33 - koronoidný proces
Lebka viacerých plazov je charakterizovaná prítomnosťou dobre definovanej časové oblúky (v jašterici sú znížené). Iba na monitore monitora vynikajúce časové oblúkyna ktorých vzdelávaní sa zúčastňujú orbitálne a šupinaté kostí. Tieto oblúky sa oddeľujú postranné časové doly, Krokodíly, spolu s hornými oblúkmi, majú dolnáktoré sa tvoria maxilárny a zygomatickýostatky (diapsidná lebka) (Obr. 52). v korytnačkyneexistujú žiadne časové jámy a časové oblúky ( lebka typu anapsid).
Viscerálna lebkapozostáva z Čeľuste a hyoidné prístroje . ostatkyvrchný čeľusť (intermaxilárne a maxilárne), ako už bolo uvedené vyššie, boli posilnené a hlboko zakorenené v mozgovej lebke. Uskutočňuje sa autostiliya - priame spojenie viscerálnej lebky s mozgom. V dôsledku osifikácie a transformácie chrupavky z palatínového štvorca štvorcový a stĺpcový kostí. Palatín a pterygoid tieto prvky sú kožného pôvodu a podieľajú sa na tvorbe spodnej časti mozgovej lebky.
Miesto má niekoľko plazov (krokodíly, korytnačky) podnebie sekundárnej kosti, ktorý vznikol rastom mnohých kostí - intermaxilárny, maxilárny, palatín a alar, Krokodíly sa zmenili a nosová dierkaVonkajšie nosné dierky sa zdvihnú nad povrch hlavy a choany sa presunú do týlnej oblasti lebky. Tieto transformácie sú základom oddelenia nosohltanu od ústnej dutiny, čo umožňuje zvieratám udržiavať svoje ústa otvorené pod vodou a súčasne dýchať cez nosné dierky vystavené povrchu vody.
Dolná čeľusť tvorené osifikáciou meckelova chrupavka a vystúpenia kĺbová, zubná, uhlová, klenutá, koronálna kosti a množstvo menších formácií. Z uvedených kostí má iba artikulárny pôvod chondrálny pôvod, všetky ostatné sú integumentary.
Sublingválne prístroje predstavuje rekord (homologická kopula) pomocou tri páry rohov - deriváty hyoidov a bočných oblúkov. Hyomandibulárne, ako u obojživelníkov, vo forme sluchovej kukly ( strmeň)umiestnené v strednom uchu.
Vybavenie a materiály
1. Kostry jašterice, korytnačky, hady (jedna súprava pre dvoch študentov).
2. Jašterica stavca z rôznych častí tela.
3. Predné a zadné končatiny jašterice s pásmi.
4. Lebka jašterice, korytnačky, hady.
5. Tabuľky: 1) kostra jašterice; 2) stavca stavcov z rôznych častí tela; 3) lebka (pohľad zhora a zdola); 4) kostra končatín a ich opasky.
Úvodné poznámky
Hlavným smerom vývoja plazov bolo zlepšenie pohybového aparátu a zvýšenie diverzity pohybov. Plazy (moderné a vyhynuté) demonštrujú všetky hlavné spôsoby pohybu, ktoré sú stavovcom známe.
V moderných jaštericiach sa zachoval evolučne originálny a typický spôsob pohybu. Rovnako ako u chvostových obojživelníkov sa pohyb vykonáva striedavým usporiadaním relatívne krátkych končatín umiestnených do strán
pri ohýbaní tela doprava - doľava. Štruktúra kĺbov končatín a všeobecný model pohybu v moderných jaštericiach sú v porovnaní s skorými plazmi iba mierne upravené.
Účinnosť tejto metódy pohybu medzi plazmi sa zvýšila v porovnaní s obojživelníkmi. To sa dosiahlo na jednej strane posilnením a diferenciáciou svalov tela a končatín a na druhej strane rozlíšením kostry a posilnením a transformáciou jej jednotlivých uzlov a kĺbov. Posilnenie svalov končatín, rozvoj pozdĺžnych svalov tela, posun končatín v jednom alebo druhom rozsahu pod telom - to všetko umožnilo rýchly pohyb s telom stúpajúcim nad zemou (beh). Posilnenie svalov a pohyb v rôznom prostredí bolo sprevádzané zvýšením zaťaženia kostry. Vyžaduje si to posilnenie. Preto hrubá vláknitá tkanina rúrkové kosti zmenené na jemne vláknité, kľúčové kosti začali hrať úlohu medzier medzi dorzálnymi (distálnymi okrajmi lopatky) a ventrálnymi (hrudnými) časťami ramenného pletenca. Najbežnejším bol percentuálny typ stavcov. Všetky stavce kmeňa, ako aj časť krčka a chvosta, nesú pohyblivé rebrá. Intenzívne pohyby flexie - predĺženie ruky a chodidla bolo možné vďaka objaveniu sa medzikarpálnych a medzistranných kĺbov.
Plazy sa vyznačujú väčšou pohyblivosťou hlavy. Toto je zabezpečené extrémne pohyblivým kĺbom lebky krčnej chrbtice chrbtica: po prvé, lebka sa spája s prvým krčným stavcom pomocou jedného kondyla; po druhé, prvé dva krčné stavce (atlas a epistrofia) sú modifikované tak, že sú možné pohyby hlavy okolo pozdĺžnej osi tela. Pohyblivosť hlavy a krku v kombinácii so schopnosťou rýchleho pohybu okolo substrátu poskytla plazom široké možnosti na nájdenie potravy. Silné stlačenie čeľustí sa ukázalo ako dôležité pri zachytávaní rôznorodej koristi. V tomto ohľade sa vyvinuli silné žuvacie svaly a prestavali sa strecha a boky lebky - objavili sa dočasné jamy, ktoré významne zväčšili povrch na pripevnenie čeľusťových svalov. Deformácia lebky pri stlačení žuvacie svaly bránené stĺpcovými kosťami.
Spojenie rebier s hrudnou kosťou a formovanie hrudníka je ďalšou významnou transformáciou kostry plazov. V dôsledku toho prudko vzrástla rýchlosť ventilácie pľúc a zvýšil sa objem čerpaného vzduchu. To prispelo k odmietnutiu dýchania kože a zvýšeniu intenzity výmeny plynov.
Kostra plazov má vo všeobecnosti komplex vlastností, ktoré preukazujú vyššiu úroveň organizácie týchto zvierat a hlbšie prispôsobenie sa životu na súši.
Preskúmajte kostru plazov.
brať do úvahy:
lebka
Mozgová lebka
tylový: začiatok; hlavné a dve bočné týlne kosti; týlny foramen; týlny kondyl.
Strany lebky: Malar; šupinatý; plávacie blanou; bar; štvorec; ušné kosti.
Strecha lebky: nosové; prefrontálnej; čelné; parietálnej; interparietal; roztrhnutie; čelo; postorbitálne kosti.
Dno lebky: hlavná sfenoidná kosť; otvárač; základy sekundárneho kostného poschodia.
Viscerálna lebka
Horná čeľusť: čeľustnej; intermaxilárne kosti.
Dolná čeľusť: zubné; kĺbovej; uhol; klenuté kosti;
Sublingválne prístroje: hyoidná kosť s dvoma pármi rohov.
chrbtica
Chrbtica: krku; prsníka; bedrový; sakrálne; chvost; stavca kufra; atlanta; epifora. hrudník: hrudnej kosti; rebrá.
Koncový pás
Ramenné pásy: hrudnej kosti; hrudný hrot; coracoid; lopatky; kľúčnu kosť.
Pelvický opasok: bedrové; sedacieho; pubické kosti; acetabulum.
Párové končatiny
predné končatiny: rameno; predlaktie (ulna a polomer); zápästie; sponka; prsty na prstoch; medzikarpálny kĺb.
Zadná končatina: stehná; palička (holenná a holenná); tarsus; priehlavok; prsty na prstoch; medzistranný kĺb.
skica:
1) lebka (pohľad zhora, pohľad zboku, dole); 2) pás predných končatín; 3) panvového pletenca a sakrálnej chrbtice.
Štruktúra kostry
Mozgová lebka
V jaštericiach je lebka takmer úplne osifikovaná a pozostáva z veľkého počtu kostí, ktoré tvoria dosť masívny mozgu box (Obr. 70, 71).
Týlna oblasť lebky pozostáva zo štyroch týlnych kostí, ktoré obmedzujú týlne foramen. Hlavná týlová kosť spolu s dvomi bočnými týlnymi kosťami tvorí týlny kondyl, ktorý vstupuje do kĺbovej dutiny atlasu. Horná týlna kosť v tvare oblúka zakrýva týlne labky zhora a vstupuje za strechu lebky.
Strany lebky v oblasti sluchovej kapsuly sú tvorené tromi ušnými kosťami, ktoré sa podieľajú na tvorbe časových jamiek.
Obr. 70. Štruktúra lebky jašterice:
- pohľad zhora; B - pohľad zdola; - bočný pohľad; 1
- týlny foramen; 2
- laterálna týlová kosť; 3
- vynikajúca týlová kosť; 4
- hlavná týlová kosť; 5
- týlny kondyl; 6
- hlavná sfenoidná kosť; 7
- otvárač; 8
- výber; 9
- parazenoid; 10
- parietálna kosť; 11
- otvor pre parietálny orgán; 12
- čelná kosť; 13
- nosovej kosti; 14
- intermaxilárna kosť; 15
- maxilárna kosť; 16
- prefrontálna kosť; 17
- slzná kosť; 18
- infraorbitálne kosti; 19
- postorbitálna (zadná) kosť; 20
- šupinatá kosť; 21
- časná kosť; 22
- zygomatická kosť; 23
- nosovej dierky; 24
- štvorcová kosť; 25
- palatínová kosť; 26
- pterygoidná kosť; 27
- stĺpcová kosť; 28
- priečna kosť; 29
- kĺbová kosť; 30
- zubná kosť; 31
- uhlová kosť; 32
- klenutá kosť; 33
- koronárna kosť

Obr. 71. Štruktúra lebky monitora jašterice:
- pohľad zhora B - pohľad zdola 1
- intermaxilárna kosť, 2
- nosová kosť 3
- prefrontálna kosť, 4
- čelná kosť 5
- slzná kosť 6
- infraorbitálna kosť 7
- parietálna kosť, 8
- vynikajúca týlová kosť, 9
- hlavná týlová kosť, 10
- laterálna týlová kosť, 11
- šupinatá kosť 12
- štvorcová kosť 13
- postorbitálna (zadná) kosť, 14
- zygomatická kosť, 75 - maxilárna kosť, 16
- otvárač 17
- palatínová kosť, 18
- priečna kosť 19
- pterygoidná kosť 20
- hlavná sfenoidná kosť, 21
- týlne kosti, 22
- štvorcová zygomatická kosť, 23
- týlny kondyl
Predné kosti sú dobre vyvinuté a izolované, horné aurálne kosti sa spájajú s hornou týlnou kosťou a zadné kosti s bočnými týlnymi kosťami. Po stranách lebky sú veľké oko. Interorbitálne septum je tvorené procesmi frontálnych a parietálnych kostí strechy lebky. Za obežnou dráhou sú postorbitálne, zygomatické a šupinaté kosti. Malé lakrimálne kosti sú umiestnené v prednom rohu obežnej dráhy a niekoľko malých infraorbitálnych kostí ohraničuje obežnú dráhu zhora. Predná časť lebky je tvorená derivátmi viscerálnej lebky - maxilárnych a intermaxilárnych kostí. V čuchovej oblasti jašterica nemá osifikáciu, čuchová kapsula s vonkajšou nosovou dierkou je tvorená chrupavkou.
Strecha lebky tvoria sa párové nosné, prefrontálne, frontálne a parietálne kosti. Medzi parietálnymi kosťami leží nepárová medziparietálna kosť s otvorom pre parietálny orgán.
Dno lebky tvorené hlavným sfenoidná kosťs ktorými sa primárne parasfenoidové poistky spoja. Na prednej časti parazofenoidu sú palatínové kosti a otvárače, na ktorých stranách
tam sú hoans. Niektoré kosti viscerálnej lebky sa tiež podieľajú na tvorbe spodnej časti mozgovej skrine - jedná sa o palatín a kosti pterygoidov, Zakrivené pterygoidné kosti s prednými koncami priliehajú k palatínovým kostiam a zadné konce kĺbovo s hlavným sfénoidom a hranatými kosťami.
V časovej oblasti jašteríc sa nachádza časná fossa, ohraničená jediným horným časovým oblúkom, ktorý je tvorený postorbitálnymi a šupinatými kosťami. Dolný časový oblúk v jaštericiach je znížený. V hĺbkach spánkových fosílií sú stĺpcové (elytra) kosti zreteľne viditeľné vo forme malých vzpier priľahlých na pterygoidné a parietálne kosti.
Viscerálna lebka
Čeľusť oblúka u jašteríc je silne upravený. V dôsledku osifikácie chrupavky z palatínového štvorca vytvorili štvorcové, pterygoidné, stĺpovité (elytra okrídlené) a palatínové kosti. Štvorcová kosť s jej horným koncom je pohyblivo kĺbovo spojená so šupinatou kosťou a spodná s dolnou čeľusťou a slúži ako jej zavesenie. Vzhľad kĺbu medzi šupinatou kosťou a horným koncom štvorcovej kosti nám umožnil zvýšiť kompresnú silu čeľustí a zjavne spôsobil evolučný úspech jašteríc. Horná čeľusť tvorené maxilárnymi a intermaxilárnymi kosťami, na ktorých sú umiestnené zuby. Medzi pterygoidnými a maxilárnymi kosťami ležia priečne kosti, ktoré pôsobia ako rozpery a posilňujú lebku.
Základ pre vývoj kostí dolná čeľusť slúži meckel chrupavky. Nahrádza sa kĺbová kosť, ktorá sa kĺbovo spojuje so štvorcovou kosťou. Okrem toho dolná čeľusť obsahuje množstvo celých kosti: najväčšie zubné kosti, ktoré nesú zuby, a menšie - koronárne, hranaté a klenuté.
Horné oddelenie podjazykový arch - Hyomandibulárny - je súčasťou stredného ucha vo forme sluchovej kukly - spony. Zvyšok hyoidného oblúka a zvyšky žiabrových oblúkov sa premenia na hyoidné zariadenie, ktoré predstavuje chrupavková hyoidná platňa a tri páry rohov.
chrbtica
Chrbát plazov je rozdelený do štyroch častí: krčné, hrudné, bedrové, krížové a chvostové.
Väčšina plazov má stavce protselnye, t.j. predná konkávna. Telo stavcov je pretiahnuté, v prednej časti je konkávne,

Obr. 72. Prvé dve krčné stavce monitora jašterice:
- atlas; B - epistrofia; 1
- horný oblúk; 2
- miechový kanál; 3
4
- zubný proces epistrofie; 5
- telo stavcov
superior duta končí vynikajúcim odstredivým procesom. Predné a zadné kĺbové procesy siahajú od tela stavcov. Na spodnej časti horného oblúka sú kĺbové povrchy na pripevnenie rebier.
Prvé dve krčné stavce sa veľmi líšia od ostatných. Prvý krčný stavca - atlas - má tvar krúžku rozdeleného hustým zväzkom na hornú a dolnú polovicu (obr. 72). Na prednej strane atlasu je kĺbová dutina pre týlny kondyl. Prostredníctvom hornej diery sa mozog pripája k mieche a zubovitý proces druhého krčného stavca vstupuje do spodnej diery v chrbte - epistrofeya, Toto spojenie lebky s krkom poskytuje zložité pohyby hlavy. Zadné krčné stavce nesú rebrá.
Na stavce hrudný zúčastniť sa rebráktoré sa kĺbovo spájajú s hrudnou kosťou a tvoria rebrovú klietku (pravé rebrá). Bedrové stavce majú tiež rebrá, ktoré sa nepripájajú k hrudnej kosti (nepravé rebrá). Dva sakrálne stavce majú masívne priečne procesy, ktoré sú spojené s panvovým pletencom (pozri obr. 75). Počet stavcov v jaštericiach sa líši a dosahuje niekoľko desiatok. Predné kaudálne stavce nesú priečne a odstredivé procesy (obr. 73) a na konci chvosta sú stavce premenené na tyčinkovité kosti. Telá takmer všetkých kaudálnych stavcov sú oddelené tenkou chrupavkovou vrstvou, pozdĺž ktorej sa počas autotómie rozbije.

Obr. 73. Štruktúra predného kaudálneho stavca jašterice:
- pohľad zhora; B - bočný pohľad; 1
- telo stavca; 2
- vynikajúci odstredivý proces; 3
- predný kĺbový proces; 4
- zadný kĺbový proces; 5
- priečny proces; 6
- spodný odstredivý proces; 7
- dolný oblúk
Koncový pás
Predný pás u jašteríc (obr. 74) je spravidla usporiadaná podobne ako ramenný pás obojživelníkov, avšak v porovnaní s ňou je tu osifikácia vysoko rozvinutá. Na križovatke kokosu so lopatkou je kĺbová dutina na kĺbové spojenie s ramennej, Supraskapulárna chrupavka rastie k dorzálnemu okraju lopatky a prokokaidná chrupavka rastie pred kokosidom. Kakaoid a prokoakarid sa spoja a tvoria jediný kostrový prvok, ktorý je kĺbovo spojený s hrudnou kosťou. V tomto kĺbe sú možné posuvné pohyby v horizontálnej rovine pozdĺž predného okraja hrudnej kosti, čo zvyšuje dĺžku kroku. Zospodu k hrudnej kosti rastie krížovité suprasternum integumentary pôvodu. Klavikuly spájajú chrbtový okraj lopatky s pectoralis. Na rozdiel od obojživelníkov sú teda plazivé goliere orientované vo vertikálnej rovine a zvyšujú odolnosť ramenného pletenca.
Panvový opasok u jašteríc je dobre vyvinutá a skladá sa z dvoch anonymných kostí, ktoré vznikli fúziou panvových kostí - iliakálnej, sedacieho a ochlpenia (obr. 75). Pre hlavicu stehennej kosti (acetabulum) existujú kĺbové dutiny. Pravá a ľavá ochlpenie a ischiálne kosti sú spojené chrupavkovou symfýzou. Panvový opasok je pevne spojený s chrbticou rastom ilium k priečnym procesom sakrálnych stavcov.
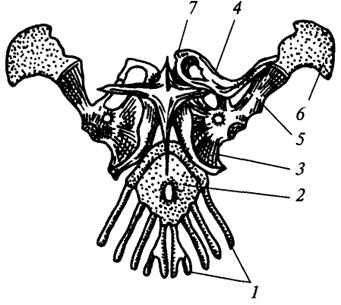
Obr. 74. Ramenný opasok jašterice:
1
- prvé štyri hrudné rebrá; 2
- hrudná kosť; 3
- kokosový; 4
- kľúčna kosť; 5
- lopatka; 6
- supraskapulárna chrupavka; 7
- hrudný pás

Obr. 75. Pelvický pás jašterice: br 1
- kĺbová (acetabulárna) dutina pre femorálnu hlavu; 2
- ilium; 3
- ochlpenie kostí; 4
- ischiálna kosť; 5
- syfýza
Párové končatiny
Končatiny plazov zodpovedajú všeobecnej štruktúre päťprstej končatiny suchozemských stavovcov. kostra predné končatiny obsahuje ramennej, ulnárne a radiálne kosti, dva rady malých kostí zápästia, rad metakarpálnych kostí a rad kostí prstov prstov. kostra zadná končatina zahŕňa stehennú kosť, holennú kosť a holennú kosť, dve rady malých tarzálnych kostí, množstvo metatarzálnych kostí a sériu prstov na prstoch. Medzi zvláštnosti štruktúry končatín patrí formácia interkarpalnogo (v prednej končatine) a invariant tertarzalnogo (v zadnej končatine) kĺbov (Obr. 76).

Obr. 76. Štruktúra medzistranného kĺbu jašterice (pravá noha rýchleho jašterica zhora):
1
- tavené proximálne kosti dechtu; 2
- distálne kosti tarzu; 3
- holenná kosť; 4
- malá holenná kosť; 5
- metatarzálne kosti
Medzikarpálny kĺb je vytvorený medzi dvoma radmi zápästových semien, medziprstný kĺb je medzi dvoma radmi kostí dechtu. Tieto kĺby poskytujú možnosť silného ohybu a roztiahnutia rúk a nôh, čo je dôležité pri zvláštnom type pohybu.