Охарактеризуйте основные типы мышечной ткани. Гистологическое строение мышечных тканей.
Мышечными тканями называют ткани, различные по строению и происхождению, но сходные по способности к выраженным сокращениям. Они обеспечивают перемещения в пространстве всего организма в целом или его частей (пример – скелетная мускулатура) и движение органов внутри орг-ма (пример – сердце, язык, кишечник).
Св-вом изменения формы обладают клетки многих тканей, но в мышечных тканях эта способность становится главной ф-цией.
Общая характеристика и классификация
Основные морфологические признаки элементов мышечных тканей - удлиненная форма, наличие продольно расположенных миофибрилл и миофиламентов - специальных органелл, обеспечивающих сократимость, расположение митохондрий рядом с сократительными элементами, наличие включений гликогена, липидов и миоглобина.
Гладкая мышечная ткань.
Эта ткань образована из мезенхимы. Клетки гладкомышечной ткани - миоциты вытянутой с заостренными концами веретеновидной формы, покрыты клеточной оболочкой. Миоцит - это одноядерная клетка. Палочковидное ядро занимает в клетке центральное положение. Оно содержит распыленный гетерохроматин и одно или два хорошо заметных ядрышка. Клеточный центр (цитоцентр) находится вблизи одной из поверхностей ядра. Остальные органеллы сконцентрированы у полюсов палочковидного ядра. Хорошо развиты митохондрии, комплекс Гольджи, гладкая ЭПС; незначительно выражены рибосомы, зернистая ЭПС. В цитоплазме миоцитов содержатся гранулы гликогена - энергетический резерв клетки.
Миоциты плотно прилегают друг к другу, образуя слои и группы, разделенные между собой рыхлой неоформленной соединительной тканью. В цитоплазме расположены миофибриллы, они идут по периферии клетки вдоль ее оси. Состоят из тонких нитей и являются сократительным элементом мышцы.
Снаружи мышечное волокно покрыто оболочкой - сарколеммой, состоящей из внутреннего и наружного слоев. Внутренний слой - это плазмолемма, которая аналогична оболочке других тканевых клеток. Наружный - соединительнотканный слой состоит из базальной мембраны и прилегающих к ней волокнистых структур. Плазмолемма образует систему узких канальцев, проникающих внутрь мышечного волокна.
Соединительнотканные волокна, расположенные снаружи базальной мембраны мышечного волокна, образуют эндомизий, который богат кровеносными сосудами и нервами. Эндомизий соединяется с перимизием - оболочкой, покрывающей группу мышечных волокон. Перимизий нескольких мышечных пучков соединяется с эпимизием - самой наружной соединительнотканной оболочкой, объединяющей несколько таких пучков в мышцу - орган, характеризующийся специфическим строением и функцией.
Встречается гладкая мышечная ткань в стенках желудка, кишечника, матки, мочевого пузыря, мочеточников, бронхов, а также в средних и крупных кровеносных сосудах. Деятельность гладких мышц регулируется ВНС. Мышечные сокращения не подчиняются воле человека и поэтому гладкую мышечную ткань называют непроизвольной мускулатурой.
Поперечнополосатая мышечная ткань.
Скелетная мышечная ткань - это сократительная ткань туловища, головы, конечностей, глотки, гортани, верхней половины пищевода, языка, жевательных мышц.
Формируется из миотомов сомитов мезодермы. Структурная единица - поперечнополосатое мышечное волокно. Имеет цилиндрическое тело, покрыто оболочкой - сарколемой, а цитоплазма называется - саркоплазмой, в которой находятся многочисленные ядра и миофибриллы. Каждая миофибрилла состоит из дисков имеющих разный химический состав. Миофибриллы являются сократительным аппаратом мышечного волокна.
Из поперечнополосатой мышечной ткани построена вся скелетная мускулатура. Мускулатура является произвольной, т.к. ее сокращение возникает под влиянием коры больших полушарий.
Мышечная ткань сердца.
Этот вид мышечной ткани образует среднюю оболочку сердца - миокард, по характеру сокращения относится к непроизвольной, так как не контролируется волей животного. Развивается она из висцерального листка спланхнатома - миоэпикардиальной пластинки.
Сердечная мышечная ткань состоит из мышечных клеток - кардиомиоцитов (сердечных миоцитов). Миоциты, соединяясь друг с другом своими концами по длинной оси клеток, формируют структуру, сходную с мышечным волокном.
Имеются два вида клеток: типичные сократительные клетки и атипичные сердечные миоциты , составляющие проводящую систему сердца.
Типичные мышечные клетки выполняют сократительную функцию; они прямоугольной формы, в центре находятся 1-2 ядра, миофибриллы расположены по периферии. Между соседними миоцитами имеются вставочные диски. С их помощью миоциты собираются в мышечные волокна, разделенные между собой тонковолокнистой соединительной тканью. Между соседними мышечными волокнами проходят соединительные волокна, которые обеспечивают сокращение миокарда, как единого целого.
Проводящая система сердца образована мышечными волокнами, состоящими из атипичных мышечных клеток . Они более крупные, чем сократительные, богаче саркоплазмой, но беднее миофибриллами, которые часто перекрещиваются. Ядра крупнее и не всегда находятся в центре. Волокна проводящей системы окружены густым сплетением нервных волокон.
Мышечными тканями (лат. textus muscularis) называют ткани, различные по строению и происхождению, но сходные по способности к выраженным сокращениям. Мышечные ткани состоят из вытянутых клеток, которые принимают раздражение от нервной системы и отвечают на него сокращением. Они обеспечивают перемещения в пространстве организма в целом, его движение органов внутри организма (сердце, язык, кишечник и др.) и состоят из мышечных волокон. Свойством изменения формы обладают клетки многих тканей, но в мышечных тканях эта способность становится главной функцией.
Основные морфологические признаки элементов мышечных тканей: удлиненная форма, наличие продольно расположенных миофибрилл и миофиламентов — специальных органелл, обеспечивающих сократимость, расположение митохондрий рядом с сократительными элементами, наличие включений гликогена, липидов и миоглобина.
Специальные сократительные органеллы — миофиламенты или миофибриллы обеспечивают сокращение, которое возникает при взаимодействии в них двух основных фибриллярных белков — актина и миозина — при обязательном участии ионов кальция. Митохондрии обеспечивают эти процессы энергией. Запас источников энергии образуют гликоген и липиды. Миоглобин — белок, обеспечивающий связывание кислорода и создание его запаса на момент сокращения мышцы, когда сдавливаются кровеносные сосуды (поступление кислорода при этом резко падает).
Свойства мышечной ткани:
- Возбудимость
- Проводимость
- Сократимость
- Лабильность
Виды мышечной ткани:
1. Гладкая мышечная ткань
Гладкая мышечная ткань состоит из одноядерных клеток — миоцитов веретеновидной формы длиной 20—500 мкм. Их цитоплазма в световом микроскопе выглядит однородно, без поперечной исчерченности. Эта мышечная ткань обладает особыми свойствами: она медленно сокращается и расслабляется, обладает автоматией, является непроизвольной (то есть ее деятельность не управляется по воле человека). Входит в состав стенок внутренних органов: кровеносных и лимфатических сосудов, мочевыводящих путей, пищеварительного тракта (сокращение стенок желудка и кишечника).
2. Поперечно-полосатая скелетная мышечная ткань
Состоит из миоцитов, имеющих большую длину (до нескольких сантиметров) и диаметр 50—100 мкм; эти клетки многоядерные, содержат до 100 и более ядер; в световом микроскопе цитоплазма выглядит как чередование тёмных и светлых полосок. Свойствами этой мышечной ткани является высокая скорость сокращения, расслабления и произвольность (то есть её деятельность управляется по воле человека). Эта мышечная ткань входит в состав скелетных мышц, а также стенки глотки, верхней части пищевода, ею образован язык, глазодвигательные мышцы. Волокна длиной от 10 до 12 см.
3. Поперечно-полосатая сердечная мышечная ткань
Состоит из 1 или 2-х ядерных кардиомиоцитов, имеющих поперечную исчерченность цитоплазмы(по периферии цитолеммы). Кардиомиоциты разветвлены и образуют между собой соединения — вставочные диски, в которых объединяется их цитоплазма.Существует также другой межклеточный контакт- аностамозы(впячивание цитолеммы одной клетки в цитолемму другой) Этот вид мышечной ткани образует миокард сердца. Развивается из миоэпикардальной пластинки (висцерального листка спланхнотома шеи зародыша) Особым свойством этой ткани является автоматия — способность ритмично сокращаться и расслабляться под действием возбуждения, возникающего в самих клетках(типичные кардиомиоциты). Эта ткань является непроизвольной(атипичные кардиомиоциты). Существует 3-й вид кардиомиоцитов- секреторные кардиомиоциты (в них нет фибрилл) Они синтезируют гормон тропонин, понижающий АД и расширяющий стенки кровеносных сосудов.
Мышечная оболочка сердца (миокард) - образована мышечными полостными клетками, которые с помощью вставочных дисков соеденяются в мышечные комплексы или сердечные мышечные волокна. Мышечная ткань имеет свойства восстанавливаться, она защищена соеденительной тканью, образующий рубец.
Отправить свою хорошую работу в базу знаний просто. Используйте форму, расположенную ниже
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
Размещено на http://www.allbest.ru/
Мышечные ткани объединяет способность к сокращению.
Особенности строения: сократительный аппарат, занимающий значительную часть в цитоплазме структурных элементов мышечной ткани и состоящий из актиновых и миозиновых филаментов, которые формируют органеллы специального назначения - миофибриллы .
Мышечные ткани представляют собой группу тканей различного происхождения и строения, объединенных на основании общего признака - выраженной сократительной способности, благодаря которой они могут выполнять свою основную функцию - перемещать тело или его части в пространстве.
Важнейшие свойства мышечных тканей. Структурные элементы мышечных тканей (клетки, волокна) обладают удлиненной формой и способны к сокращению благодаря мощному развитию сократительного аппарата. Для последнего характерно высокоупорядоченное расположение актиновых и миозиновых миофиламентов, создающее оптимальные условия для их взаимодействия. Это достигается связью сократимых структур с особыми элементами цитоскелета и плазмолеммой (сарколеммой), выполняющими опорную функцию. В части мышечных тканей миофиламенты образуют органеллы специального значения - миофибриллы. Для мышечного сокращения требуется значительное количество энергии, поэтому в структурных элементах мышечных тканей имеется большое количество митохондрий и трофических включений (липидных капель, гранул гликогена), содержащих субстраты - источники энергии. Поскольку мышечное сокращение протекает с участием ионов кальция, в мышечных клетках и волокнах хорошо развиты структуры, осуществляющие его накопление и выделение - агранулярная эндоплазматическая сеть (саркоплазматическая сеть), кавеолы.
Классификация мышечных тканей основана на признаках их (а) строения и функции (морфофункциональная классификация) и (б) происхождения (гистогенетическая классификация).
Морфофункциональная классификация мышечных тканей выделяет поперечнополосатые (исчерченные) мышечные ткани и гладкую мышечную ткань. Поперечнополосатые мышечные ткани образованы структурными элементами (клетками, волокнами), которые обладают поперечной исчерченностью вследствие особого упорядоченного взаиморасположения в них актиновых и миозиновых миофиламентов. К поперечнополосатым мышечным тканям относят скелетную и сердечную мышечную ткани. Гладкая мышечная ткань состоит из клеток, не обладающих поперечной исчерченностью. Наиболее распространенным видом этой ткани является гладкая мышечная ткань, входящая в состав стенки различных органов (бронхов, желудка, кишки, матки, маточной трубы, мочеточника, мочевого пузыря и сосудов).
Гистогенетическая классификация мышечных тканей выделяет три основных типа мышечных тканей: соматический (скелетная мышечная ткань), целомический (сердечная мышечная ткань) и мезенхимный (гладкая мышечная ткань внутренних органов), а также два дополнительных: миоэпителиальные клетки (видоизмененные эпителиальные сократимые клетки в концевых отделах и мелких выводных протоках некоторых желез) и мионейральные элементы (сократимые клетки нейрального происхождения в радужке глаза).
Скелетная поперечнополосатая (исчерченная) мышечная ткань по своей массе превышает любую другую ткань организма и является самой распространенной мышечной тканью тела человека. Она обеспечивает перемещение тела и его частей в пространстве и поддержание позы (входит в состав локомоторного аппарата), образует глазодвигательные мышцы, мышцы стенки полости рта, языка, глотки, гортани. Аналогичное строение имеет нескелетная висцеральная исчерченная мышечная ткань, которая обнаруживается в верхней трети пищевода, входит в состав наружных анального и уретрального сфинктеров.
мышечный миоцит сердечный скелетный
Скелетная поперечнополосатая мышечная ткань развивается в эмбриональном периоде из миотомов сомитов, дающих начало активно делящимся миобластам - клеткам, которые располагаются цепочками и сливаются друг с другом в области концов с образованием мышечных трубочек (миотубул) , превращающихся в мышечные волокна. Такие структуры, образованные единой гигантской цитоплазмой и многочисленными ядрами, в отечественной литературе традиционно именуют симпластами (в данном случае - миосимпластами), однако этот термин отсутствует в принятой международной терминологии. Некоторые миобласты не сливаются с другими, располагаясь на поверхности волокон и давая начало миосателлитоцитам - мелким клеткам, которые являются камбиальными элементами скелетной мышечной ткани. Скелетная мышечная ткань образована собранными в пучки поперечнополосатыми мышечными волокнами , являющимися ее структурно-функциональными единицами.
Мышечные волокна скелетной мышечной ткани представляют собой цилиндрические образования вариабельной длины (от миллиметров до 10-30 см). Их диаметр также широко варьирует в зависимости от принадлежности к определенной мышце и типу, функционального состояния, степени функциональной нагрузки, состояния питания и других факторов. В мышцах мышечные волокна образуют пучки, в которых они лежат параллельно и, деформируя друг друга, часто приобретают неправильную многогранную форму, что особенно хорошо видно на поперечных срезах. Между мышечными волокнами располагаются тонкие прослойки рыхлой волокнистой соединительной ткани, несущие сосуды и нервы - эндомизий. Поперечная исчерченность скелетных мышечных волокон обусловлена чередованием темных анизотропных дисков (полос А) и светлых изотропных дисков (полос I). Каждый изотропный диск рассекается надвое тонкой темной линией Z - телофрагмой . Ядра мышечного волокна - сравнительно светлые, с 1-2 ядрышками, диплоидные, овальные, уплощенные - лежат на его периферии под сарколеммой и располагаются вдоль волокна. Снаружи сарколемма покрыта толстой базальной мембраной, в которую вплетаются ретикулярные волокна.
Миосателлитоциты (клетки-миосателлиты) - мелкие уплощенные клетки, располагающиеся в неглубоких вдавлениях сарколеммы мышечного волокна и покрытые общей базальной мембраной (см. рис. 88). Ядро миосателлитоцита - плотное, относительно крупное, органеллы мелкие и немногочисленные. Эти клетки активируются при повреждении мышечных волокон и обеспечивают их репаративную регенерацию. Сливаясь с остальной частью волокна при усиленной нагрузке, миосателлитоциты участвуют в его гипертрофии.
Миофибриллы образуют сократительный аппарат мышечного волокна, располагаются в саркоплазме по ее длине, занимая центральную часть, и отчетливо выявляются на поперечных срезах волокон в виде мелких точек.
Миофибриллы обладают собственной поперечной исчерченностью, причем в мышечном волокне они располагаются столь упорядоченно, что изотропные и анизотропные диски разных миофибрилл совпадают между собой, обусловливая поперечную исчерченность всего волокна. Каждая миофибрилла образована тысячами повторяющихся последовательно связанных между собой структур - саркомеров.
Саркомер (миомер) является структурно-функциональной единицей миофибриллы и представляет собой ее участок, расположенный между двумя телофрагмами (линиями Z). Он включает анизотропный диск и две половины изотропных дисков - по одной половине с каждой стороны. Саркомер образован упорядоченной системой толстых (миозиновых) и тонких (актиновьх) миофиламентов. Толстые миофиламенты связаны с мезофрагмой (линией М) и сосредоточены в анизотропном диске,
а тонкие миофиламенты прикреплены к телофрагмам (линиям Z), образуют изотропные диски и частично проникают в анизотропный диск между толстыми нитями вплоть до светлой полосы Н в центре анизотропного диска.
В мышечной, как в других тканях, различают два вида регенерации - физиологическую и репаративную. Физиологическая регенерация проявляется в форме гипертрофии мышечных волокон, что выражается в увеличении их толщины и даже длины, увеличение числа органелл, главным образом миофибрилл, а также нарастании числа ядер, что в конечном счете проявляется увеличением функциональной способности мышечного волокна. Радиоизотопным методом установлено, что увеличение числа ядер в мышечных волокнах в условиях гипертрофии достигается за счет деления клеток миосателлитов и последующего вхождения в миосимпласт дочерних клеток.
Увеличение числа миофибрилл осуществляется посредством синтеза актиновых и миозиновых белков свободными рибосомами и последующей сборки этих белков в актиновые и миозиновые миофиламенты параллельно с соответствующими филаментами саркомеров. В результате этого вначале происходит утолщение миофибрилл, а затем их расщепление и образование дочерних миофибрилл. Кроме того возможно образование новых актиновых и миозиновых миофиламентов не параллельно, а встык предшествующим миофибриллам, чем достигается их удлинение. Саркоплазматическая сеть и Т-канальцы в гипертрофирующемся волокне образуются за счет разрастания предшествующих элементов. При определенных видах мышечной тренировки может формироваться преимущественно красный тип мышечных волокон (у стайеров) или белый тип мышечных волокон (у спринтеров). Возрастная гипертрофия мышечных волокон интенсивно проявляется с началом двигательной активности организма (1-2 года), что обусловлено прежде всего усилением нервной стимуляции. В старческом возрасте, а также в условиях малой мышечной нагрузки наступает атрофия специальных и общих органелл, истончение мышечных волокон и снижение их функциональной способности.
Репаративная регенерация развивается после повреждения мышечных волокон. При этом способ регенерации зависит от величины дефекта. При значительных повреждениях на протяжении мышечного волокна миосателлиты в области повреждения и в прилежащих участках растормаживаются, усиленно пролиферируют, а затем мигрируют в область дефекта мышечного волокна, где выстраиваются в цепочки, формируя миотрубку. Последующая дифференцировка миотрубки приводит к восполнению дефекта и восстановлению целостности мышечного волокна. В условиях небольшого дефекта мышечного волокна на его концах, за счет регенерации внутриклеточных органелл, образуются мышечные почки, которые растут навстречу друг другу, а затем сливаются, приводя к закрытию дефекта. Однако, репаративная регенерация и восстановление целостности мышечных волокон могут осуществляться при определенных условиях: во-первых, при сохраненной двигательной иннервации мышечных волокон, во-вторых, если в область повреждения не попадают элементы соединительной ткани (фибробласты). Иначе на месте дефекта мышечного волокна развивается соединительно-тканный рубец.
Советским ученым А.Н. Студитским доказана возможность аутотрансплантации скелетной мышечной ткани и даже целых мышц при соблюдении определенных условий:
· механическое измельчение мышечной ткани трансплантата, с целью растормаживания клеток-сателлитов и последующей их пролиферации;
· помещение измельченной ткани в фасциальное ложе;
· подшивание двигательного нервного волокна к измельченному трансплантату;
· наличие сократительных движений мышц-антагонистов и синергистов.
Анатомически у новорожденных имеются все скелетные мышцы, но относительно веса тела они составляют всего 23% (у взрослого 44%). Количество мышечных волокон в мышцах такое же как у взрослого. Однако микроструктура мышечных волокон отличается: волокна меньше диаметром, в них больше ядер. По мере роста происходит утолщение и удлинение волокон. Это происходит за счет утолщения миофибрилл, оттесняющих ядра на периферию. Размеры мышечных волокон стабилизируются к 20 годам.
Мышцы у детей эластичнее, чем у взрослых. Т.е. быстрее укорачиваются при сокращении и удлиняются при расслаблении. Возбудимость и лабильность мышц новорожденных, ниже, чем взрослых, но с возрастом растет. У новорожденных даже во сне мышцы находятся в состоянии тонуса. Развитие различных групп мышц происходит неравномерно. В 4-5 лет более развиты мышцы предплечья, отстают в развитии мышцы кисти. Ускоренное согревание мышц кисти происходит в 6-7 лет. Причем разгибатели развиваются медленнее сгибателей. С возрастом изменяется соотношение тонуса мышц. В раннем детстве повышен тонус мышц кисти, разгибателей бедра т.д. постепенно распределение тонуса нормализуется.
Для сердца как органа характерна способность к регенерации путем регенераторной гипертрофии, при которой масса органа восстанавливается, но форма остается нарушенной. Подобное явление наблюдается после перенесенного инфаркта миокарда, когда масса сердца может восстановиться как целое, при этом на месте повреждения образуется соединительнотканный рубец, но орган гипертрофируется, т.е. нарушается форма. Происходит не только увеличение размеров кардиомиоцитов, но и пролиферация в основном в предсердиях и ушках сердца.
Ранее полагали, что дифференцировка кардиомиоцитов является необратимым процессом, связанным с полной потерей этими клетками способности к делению. Но на современном уровне многочисленные данные показывают то, что дифференцированные кардиомиоциты способны к синтезу ДНК и митозу. В исследовательских работах П.П. Румянцева и его учеников показано, что после экспериментального инфаркта миокарда левого желудочка сердца в клеточный цикл возвращается 60-70% предсердных кардиомиоцитов, возрастает число полиплоидных клеток, но это не компенсирует повреждение миокарда.
Установлено, что кардиомиоциты способны к митотическому делению (в том числе и клетки проводящей системы). В миокарде сердца особенно много одноядерных полиплоидных клеток с 16-32-кратным содержанием ДНК, но встречаются и двуядерные кардиомиоциты (13-14%) в основном октоплоидные.
В процессе регенерации сердечной мышечной ткани кардиомиоциты участвуют в процессе гиперплазии и гипертрофии, возрастает их плоидность, но уровень пролиферации клеток соединительной ткани в области повреждения оказывается в 20-40 раз выше. В фибробластах активизируется синтез коллагена, в результате чего репарация происходит путем рубцевания дефекта. Биологическое представление подобной адаптационной реакции соединительной ткани объясняется жизненной важностью сердечного органа, так как задержка с закрытием дефекта может привести к гибели.
Считалось, что у новорожденных, а возможно, и в раннем детском возрасте, когда способные к делению кардиомиоциты еще сохраняются, регенераторные процессы сопровождаются увеличением количества кардиомиоцитов. При этом у взрослых физиологическая регенерация осуществляется в миокарде в основном путем внутриклеточной регенерации, без увеличения количества клеток, т.е. в миокарде взрослого человека отсутствует пролиферация кардиомиоцитов. Но недавно получены данные о том, что в здоровом сердце человека 14 миоцитов из миллиона находится в состоянии митоза, завершающегося цитотомией, т.е. количество клеток не значительно, но увеличивается.
Применение современных методов клеточной биологии в клинических и экспериментальных исследованиях позволило перейти к выяснению клеточных и молекулярных механизмов повреждения и регенерации миокарда. Особенно интересны данные о том, что в перинекротических областях и в функционально перегруженном сердце происходит синтез эмбриональных миоакрдиальных белков и пептидов, а также белков, синтезирующихся во время клеточного цикла. Это подтверждает положение о сходстве механизмов регенерации и нормального онтогенеза.
Выяснилось также и то, что дифференцированные кардиомиоциты в культуре способны к активному митотическому делению, что, возможно, объясняется не полной утратой, а подавлением способности кардиомиоцитов возвращаться в клеточный цикл.
Важной задачей теоретической и практической кардиологии является разработка способов стимуляции восстановления поврежденного миокарда, т.е. индукции миокардиальной регенерации и уменьшения соединительнотканного рубца. Одно из направлений исследований предоставляет возможность переноса регуляторных генов, которые превращают фибробласты рубца в миобласты или трансфекция в кардиомиоциты генов, контролирующих рост новых клеток. Другим направлением является перенос в область повреждения фетальных скелетных и миокардиальных клеток, которые могли бы участвовать в восстановлении сердечной мышцы. Также проводятся эксперименты по трансплантации скелетной мышцы в сердце, показывающие образование в миокарде участков сокращающейся ткани и улучшающие функциональные показатели миокарда. Перспективным может быть лечение с применением факторов роста, оказывающих как прямое, так и косвенное воздействие на поврежденный миокард, например, улучшение ангиогенеза.
Гладкие мышечные ткани
По происхождению различают три группы гладких (или неисчерченных) мышечных тканей -- мезенхимные, эпидермальные и нейральные.
Мышечная ткань мезенхимного происхождения
Гистогенез. Стволовые клетки и клетки-предшественники гладкой мышечной ткани, будучи уже детерминированными, мигрируют к местам закладки органов. Дифференцируясь, они синтезируют компоненты матрикса и коллаген базальной мембраны, а также эластин. У дефинитивных клеток (миоцитов) синтетическая способность снижена, но не исчезает полностью.
Структурно-функциональной единицей гладкой, или неисчерченной, мышечной ткани является гладкомышечная клетка, или гладкий миоцит -- это веретеновидная клетка длиной 20--500 мкм, шириной 5--8 мкм. Ядро клетки палочковидное, находится в ее центральной части. Когда миоцит сокращается, его ядро изгибается и даже закручивается. Органеллы общего значения, среди которых много митохондрий, сосредоточены в цитоплазме около полюсов ядра. Аппарат Гольджи и гранулярная эндо плазматическая сеть развиты слабо, что свидетельствует о малой активности синтетических функций. Рибосомы в большинстве своем расположены свободно.
Филаменты актина образуют в цитоплазме трехмерную сеть, вытянутую преимущественно продольно, точнее косо-продольно. Концы филаментов скреплены между собой и с плазмолеммой специальными сшивающими белками. Эти участки хорошо видны на электронных микрофотографиях как плотные тельца.
Миозиновые филаменты находятся в деполимеризованном состоянии. Мономеры миозина располагаются рядом с филаментами актина. Сигнал к сокращению обычно поступает по нервным волокнам. Медиатор, который выделяется из их терминалей, изменяет состояние плазмолеммы. Она образует впячивания -- кавеолы, в которых концентрируются ионы кальция. Кавеолы отшнуровываются в сторону цитоплазмы в виде пузырьков (здесь из пузырьков освобождается кальций). Это влечет за собой как полимеризацию миозина, так и взаимодействие миозина с актином. Актиновые филаменты смещаются друг другу навстречу, плотные пятна сближаются, усилие передается на плазмолемму, и вся клетка укорачивается. Когда поступление сигналов со стороны нервной системы прекращается, ионы кальция эвакуируются из кавеол, миозин деполимеризуется и «миофибриллы» распадаются. Таким образом, актино-миозиновые комплексы существуют в гладких миоцитах только в период сокращения.
Гладкие миоциты располагаются без заметных межклеточных пространств и разделены базальной мембраной. На отдельных участках в ней образуются «окна», поэтому плазмолеммы соседних миоцитов сближаются. Здесь формируются нексусы, и между клетками возникают не только механические, но и метаболические связи. Поверх «чехликов» из базальной мембраны между миоцитами проходят эластические и ретикулярные волокна, объединяющие клетки в единый тканевой комплекс. Ретикулярные волокна проникают в щели на концах миоцитов, закрепляются там и передают усилие сокращения клетки всему их объединению.
Регенерация. Физиологическая регенерация гладкой мышечной ткани проявляется в условиях повышенных функциональных нагрузок. Наиболее отчетливо это видно в мышечной оболочке матки при беременности. Такая регенерация осуществляется не столько на тканевом, сколько на клеточном уровне: миоциты растут, в цитоплазме активизируются синтетические процессы, количество миофиламентов увеличивается (рабочая гипертрофия клеток). Не исключена, однако, и пролиферация клеток (т.е. гиперплазия).
В составе органов миоциты объединяются в пучки, между которыми располагаются тонкие прослойки соединительной ткани. В эти прослойки вплетаются ретикулярные и эластические волокна, окружающие миоциты. В прослойках проходят кровеносные сосуды и нервные волокна. Терминали последних оканчиваются не непосредственно на миоцитах, а между ними. Поэтому после поступления нервного импульса медиатор распространяется диффузно, возбуждая сразу многие клетки. Гладкая мышечная ткань мезенхимного происхождения представлена главным образом в стенках кровеносных сосудов и многих трубчатых внутренних органов, а также образует отдельные мелкие мышцы.
Гладкая мышечная ткань в составе конкретных органов имеет неодинаковые функциональные свойства. Это обусловлено тем, что на поверхности органов имеются разные рецепторы к конкретным биологически активным веществам. Поэтому и на многие лекарственные препараты их реакция неодинакова.
Гладкая мышечная ткань эпидермального происхождения
Миоэпителиальные клетки развиваются из эпидермального зачатка. Они встречаются в потовых, молочных, слюнных и слезных железах и имеют общих предшественников с железистыми секреторными клетками. Миоэпителиальные клетки непосредственно прилежат к собственно эпителиальным и имеют общую с ними базальную мембрану. При регенерации те и другие клетки восстанавливаются из общих малодифференцированных предшественников. Большинство миоэпителиальных клеток имеют звездчатую форму. Эти клетки нередко называют корзинчатыми: их отростки охватывают концевые отделы и мелкие протоки желез. В теле клетки располагаются ядро и органеллы общего значения, а в отростках -- сократительный аппарат, организованный, как и в клетках мышечной ткани мезенхимного типа.
Гладкая мышечная ткань нейрального происхождения
Миоциты этой ткани развиваются из клеток нейрального зачатка в составе внутренней стенки глазного бокала. Тела этих клеток располагаются в эпителии задней поверхности радужки. Каждая из них имеет отросток, который направляется в толщу радужки и ложится параллельно ее поверхности. В отростке находится сократительный аппарат, организованный так же, как и во всех гладких миоцитах. В зависимости от направления отростков (перпендикулярно или параллельно краю зрачка) миоциты образуют две мышцы -- суживающую и расширяющую зрачок.
Заключение
Как уже было отмечено, мышечные ткани - это группа тканей организма различного происхождения, объединяемых по признаку сократимости: поперечнополосатая (скелетная и сердечная), гладкая, а также специализированные сократимые ткани - эпителиально-мышечная и нейроглиальная, входящая в состав радужки глаза.
Поперечнополосатая скелетная мышечная ткань возникает из миотомов, входящих в состав элементов сегментированной мезодермы - сомитов.
Гладкая мышечная ткань человека и позвоночных животных развивается в составе производных мезенхимы, так же как и ткани внутренней среды. Однако для всех мышечных тканей характерно сходное обособление в составе эмбрионального зачатка в виде клеток веретенообразной формы - мышцеобразовательных клеток, или миобластов.
Сокращение мышечного волокна заключается в укорочении миофибрилл в пределах каждого саркомера. Толстые (миозиновые) и тонкие (актиновые) нити, в расслабленном состоянии связанные только концевыми отделами, в момент сокращения осуществляют скользящие движения навстречу друг другу. Выделение необходимой для сокращения энергии происходит в результате превращения АТФ в АДФ под влиянием миозина. Ферментная активность миозина проявляется при условии оптимального содержания Са2+, которые накапливаются в саркоплазматической сети.
Список литературы
1. Гистология. Под редакцией Ю.И. Афанасьевой, Н.А. Юриной. М.: “Медицина”, 1999 г.
2. Р. Эккерт, Д. Рендел, Дж. Огастин “Физиология животных” - 1 т. М.: “Мир”, 1981 г.
3. К.П. Рябов “Гистология с основами эмбриологии” Минск: “Высшая школа”, 1990 г.
4. Гистология. Под редакцией Улумбекова, проф. Ю.А. Челышева. М.: 1998 г.
5. Гистология. Под редакцией В.Г. Елисеева. М.: “Медицина”, 1983 г.
Размещено на Allbest.ru
...Подобные документы
Структурные особенности мышечных тканей. Изучение механизма мышечного сокращения и аппарата передачи возбуждения. Гистогенез и регенерация мышечной ткани. Принципы работы сократительных, проводящих и секреторных кардиомиоцитов сердечной мышечной ткани.
шпаргалка , добавлен 14.11.2010
Изучение видов и функций различных тканей человека. Задачи науки гистологии, которая изучает строение тканей живых организмов. Особенности строения эпителиальной, нервной, мышечной ткани и тканей внутренней среды (соединительной, скелетной и жидкой).
презентация , добавлен 08.11.2013
Изучение особенностей строения тканей животных, функционирование и разновидности. Проведение исследования характерной черты строения соединительной и нервной тканей. Структура плоской, кубической, мерцательной и железистой эпителии. Виды мышечной ткани.
презентация , добавлен 08.02.2015
Общая характеристика и возрастные особенности хрящевой ткани. Виды хрящевой и костной ткани. Общая характеристика и возрастные особенности костной ткани. Особенности строения мышечной ткани в детском и в пожилом возрасте. Скелетная мышечная ткань.
презентация , добавлен 07.02.2016
Класификация тканей, виды эпителиальных тканей, их строение и функции. Опорная, трофическая и защитная функция соединительных тканей. Функции нервной и мышечной тканей. Понятие об органах и системах органов, их индивидуальные, половые, возрастные отличия.
реферат , добавлен 11.09.2009
Эпителиальная ткань, ее регенерационная способность. Соединительные ткани, участвующие в поддержании гомеостаза внутренней среды. Клетки кровы и лимфы. Поперечнополосатые и сердечные мышечные ткани. Функции нервных клеток и тканей животных организмов.
реферат , добавлен 16.01.2015
Изучение видов тканей животных, а также функций, которые они выполняют. Особенности строения эпителиальной, соединительной, мышечной и нервной группы тканей. Определение месторасположения каждой группы и значения для жизнедеятельности организма животного.
презентация , добавлен 18.10.2013
Общее понятие и разновидности колебаний. Характеристика процессов растяжения (сжатия), сдвига, изгиба, кручения. Механические свойства костной и сосудистой тканей. Специфика мышечной ткани, основные режимы работы мышц – изометрический и изотонический.
контрольная работа , добавлен 19.03.2014
Клетка как основная структурная единица организма. Описание ее строения, жизненных и химических свойств. Строение и функции эпителиальной и соединительной, мышечной и нервной тканей. Органы и перечень системы органов человека, их назначение и функции.
презентация , добавлен 19.04.2012
Физиология и биохимия мышечной деятельности как важная составляющая обмена веществ в организме. Типы мышечной ткани и соответственно мышц, различающихся по структуре мышечных волокон, характеру иннервации. Влияние физических нагрузок разной интенсивности.
Глава 9. МЫШЕЧНЫЕ ТКАНИ
Глава 9. МЫШЕЧНЫЕ ТКАНИ
Мышечными тканями (textus muscularis) называют ткани, различные по строению и происхождению, но сходные по способности к сокращениям. Они обеспечивают перемещения в пространстве организма в целом, его частей и движение органов внутри организма (сердце, язык, кишечник и др.).
Свойством сокращаться с изменением формы обладают клетки многих тканей, но в мышечных тканях эта способность становится главной функцией.
9.1. ОБЩАЯ МОРФОФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА И КЛАССИФИКАЦИЯ
Основные морфологические признаки элементов мышечных тканей - удлиненная форма, наличие продольно расположенных миофибрилл и миофиламентов - специальных органелл, обеспечивающих сократимость, расположение митохондрий рядом с сократительными элементами, наличие включений гликогена, липидов и миоглобина.
Специальные сократительные органеллы - миофиламенты или миофи-бриллы, обеспечивают сокращение, которое возникает при взаимодействии в них двух основных фибриллярных белков - актина и миозина, при обязательном участии ионов кальция. Митохондрии обеспечивают эти процессы энергией. Запас источников энергии образуют гликоген и липиды. Миоглобин - белок, обеспечивающий связывание кислорода и создание его запаса на момент сокращения мышцы, когда сдавливаются кровеносные сосуды (поступление кислорода при этом резко снижается).
Классификация. В основу классификации мышечных тканей положены два принципа - морфофункциональный и гистогенетический. В соответствии с морфофункциональным принципом, в зависимости от структуры органелл сокращения, мышечные ткани подразделяют на две подгруппы.
Первая подгруппа - поперечнополосатые (исчерченные) мышечные ткани (textus muscularis striatus). В цитоплазме их элементов миозиновые филамен-
ты постоянно полимеризованы, образуют с актиновыми нитями постоянно существующие миофибриллы. Последние организованы в характерные комплексы - саркомеры. В соседних миофибриллах структурные субъединицы саркомеров расположены на одном уровне и создают поперечную исчер-ченность.
Вторая подгруппа - гладкие (неисчерченные) мышечные ткани (textus muscularis nonstriatus). Эти ткани характеризуются тем, что вне сокращения миозиновые филаменты деполимеризованы. В присутствии ионов кальция они полимеризуются и вступают во взаимодействие с филаментами актина. Образующиеся при этом миофибриллы не имеют поперечной исчерченности: при специальных окрасках они представлены равномерно окрашенными по всей длине (гладкими) нитями.
В соответствии с гистогенетическим принципом в зависимости от источников развития (эмбриональных зачатков) мышечные ткани и мышечные элементы подразделяются на: соматические (миотомные), целомические (из миоэпикардиальной пластинки висцерального листка спланхнотома), мезенхимные (из десмального зачатка в составе мезенхимы), нейральные (из нервной трубки), эпидермальные (из кожной эктодермы и из прехор-дальной пластинки).
9.2. ПОПЕРЕЧНОПОЛОСАТЫЕ МЫШЕЧНЫЕ ТКАНИ
Имеется две основные разновидности поперечнополосатых (исчерченных) тканей - скелетная (миотомная) и сердечная (целомическая).
9.2.1. Скелетная мышечная ткань
Гистогенез. Источником развития элементов скелетной (соматической) поперечнополосатой мышечной ткани (textus muscularis striatus sceletalis) являются стволовые клетки миотомов - промиобласты. Одни из них дифференцируются на месте и участвуют в образовании так называемых аутохтон-ных мышц. Другие клетки мигрируют из миотомов в мезенхиму. Они уже детерминированы, хотя внешне не отличаются от других клеток мезенхимы. Их дифференцировка продолжается в местах закладки других мышц тела. В ходе дифференцировки возникают две клеточные линии. Клетки одной из линий сливаются, образуя удлиненные симпласты - мышечные трубочки (миотубы). В них происходит дифференцировка специальных органелл - миофибрилл (рис. 9.1). В это время в миотубах отмечается хорошо развитая гранулярная эндоплазматическая сеть. Миофибриллы сначала располагаются под плазмолеммой, а затем заполняют большую часть миотубы. Ядра, напротив, из центральных отделов смещаются к периферии. Клеточные центры и микротрубочки при этом полностью исчезают. Гранулярная эндо-
Рис. 9.1. Гистогенез скелетной мышечной ткани (по А. А. Клишову):
а - промиобласты; б - миосимпласт; в - мышечная трубочка; г - зрелое мышечное
волокно. 1 - миосателлитоцит; 2 - ядро миосимпласта; 3 - миофибриллы
плазматическая сеть в значительной степени редуцируется. Такие дефинитивные структуры называют миосимпластами.
Клетки другой линии остаются самостоятельными и дифференцируются в миосателлитоциты. Эти клетки располагаются на поверхности миосимпластов. Миосателлитоциты, размножаясь, сливаются с миосимпластами, участвуя таким образом в создании оптимального ядерно-
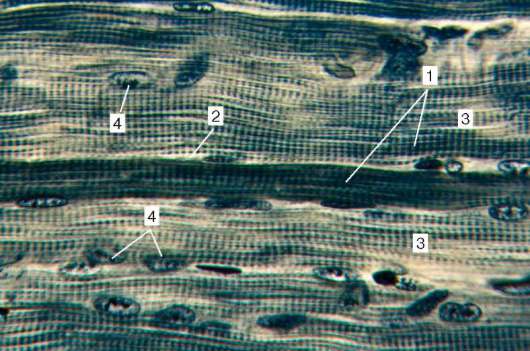
Рис. 9.2. Строение поперечнополосатой мышечной ткани (микрофотография):
1 - мышечные волокна; 2 - сарколемма; 3 - саркоплазма и миофибриллы; 4 - ядра
миосимпласта. Окраска - железный гематоксилин
саркоплазменного отношения, необходимого для синтеза специфических белков симпласта.
Строение. Основной структурной единицей скелетной мышечной ткани является мышечное волокно, состоящее из миосимпласта и миосател-литоцитов, покрытых общей базальной мембраной (рис. 9.2-9.4). Длина всего волокна может измеряться сантиметрами при толщине 50-100 мкм. Комплекс, состоящий из плазмолеммы миосимпласта и базальной мембраны, называют сарколеммой.
Строение миосимпласта. Миосимпласт имеет множество продолговатых ядер, расположенных непосредственно под плазмолеммой. Их количество в одном симпласте может достигать нескольких десятков тысяч (см. рис. 9.2). У полюсов ядер располагаются органеллы общего значения - комплекс Гольджи и небольшие фрагменты агранулярной эндоплазматической сети. Миофибриллы заполняют основную часть миосимпласта и расположены продольно (см. рис. 9.3).
Саркомер - структурная единица миофибриллы. Каждая миофибрилла имеет поперечные темные и светлые диски с неодинаковым лучепреломлением (анизотропные А-диски и изотропные I-диски). Каждая миофибрил-ла окружена продольно расположенными и анастомозирующими между собой петлями агранулярной эндоплазматической сети - саркоплазмати-ческой сети. Соседние саркомеры имеют общую пограничную структуру - Z-линию (рис. 9.5). Она построена в виде сети из белковых фибриллярных молекул, среди которых существенную роль играет альфа-актинин. С этой сетью связаны концы актиновых филаментов. От соседних Z-линий акти-новые филаменты направляются к центру саркомера, но не доходят до его середины. Филаменты актина объединены с Z-линией и нитями миозина
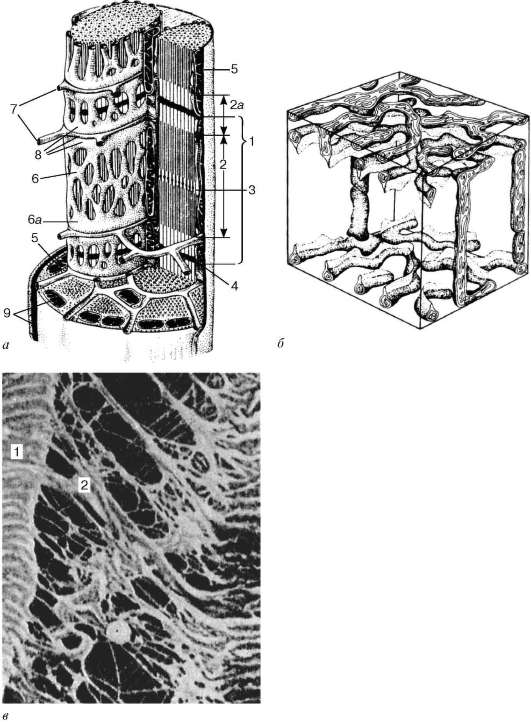
Рис. 9.3. Схема ультрамикроскопического строения миосимпласта (по Р. В. Крстичу, с изменениями) (а): 1 - саркомер; 2 - анизотропный диск (полоса А); 2а - изотропный диск (полоса I); 3 - линия М (мезофрагма) в середине анизотропного диска; 4 - линия Z (телофрагма) в середине изотропного диска; 5 - митохондрии; 6 - сар-коплазматическая сеть; 6а - конечная цистерна; 7 - поперечная трубочка (Т-трубочка); 8 - триада; 9 - сарколемма; б - схема пространственного расположения митохондрий в симпласте. Верхняя и нижняя плоскости рисунка ограничиваютанизотропныйдисксарко-мера (по Л. Е. Бакеевой, В. П. Скулачеву, Ю. С. Ченцову); в - эндомизий. Сканирующая электронная микрофотография, увеличение 2600 (препарат Ю. А. Хорошкова): 1 - мышечные волокна; 2 - коллагеновые фибриллы
фибриллярными нерастяжимыми молекулами небулина. Посередине темного диска саркомера располагается сеть, построенная из миомиозина. Она образует в сечении М-линию. В узлах этой М-линии закреплены концы мио-зиновых филаментов. Другие их концы направляются в сторону Z-линий и
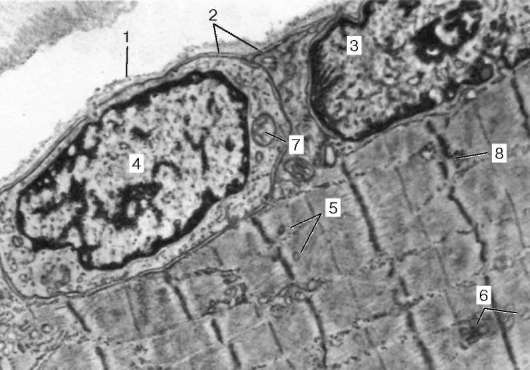
Рис. 9.4. Поверхностный участок миосимпласта и миосателлитоцит. Электронная микрофотография, увеличение 10 000 (препарат В. Л. Горячкиной и С. Л. Кузнецова): 1 - базальная мембрана; 2 - плазмолемма; 3 - ядро миосимпласта; 4 - ядро миоса-теллитоцита; 5 - миофибриллы; 6 - канальцы агранулярной эндоплазматической (саркоплазматической) сети; 7 - митохондрии; 8 - гликоген
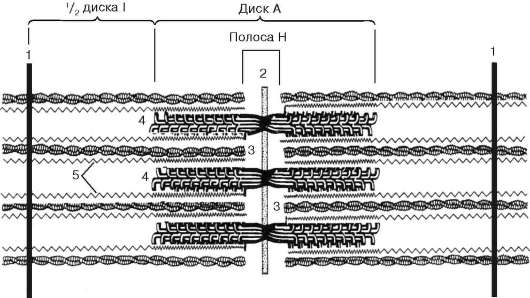
Рис. 9.5. Саркомер (схема):
1 - линия Z; 2 - линия М; 3 - филаменты актина; 4 - филаменты миозина; 5 - фибриллярные молекулы титина (по Б. Албертс, Д. Брей, Дж. Льюис и др., с изменениями)
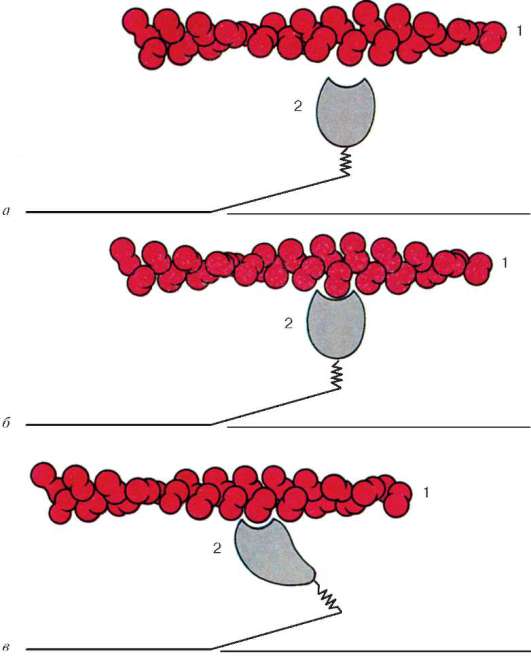
Рис. 9.6. Конформационные изменения, влекущие за собой взаимное смещение филаментов актина и миозина:
а-в - последовательные изменения пространственных отношений. 1 - актин; 2 - головка молекулы миозина (по Б. Албертс, Д. Брей, Дж. Льюис и соавт., с изменениями)
располагаются между филаментами актина, но до самих Z-линий тоже не доходят. Вместе с тем эти концы фиксированы по отношению к Z-линиям растяжимыми гигантскими белковыми молекулами титина.
Молекулы миозина имеют длинный хвост и на одном из его концов две головки. При повышении концентрации ионов кальция в области присоединения головок (шарнирный участок) молекула изменяет свою конфигурацию (рис. 9.6). При этом (поскольку между миозиновыми филаментами расположены актиновые) головки миозина связываются с актином (при участии вспомогательных белков - тропомиозина и тропонина). Затем головка миозина наклоняется и тянет за собой актиновую молекулу в сторону М-линии. Z-линии сближаются, саркомер укорачивается.
Альфа-актининовые сети Z-линий соседних миофибрилл связаны друг с другом промежуточными филаментами. Они подходят к внутренней поверхности плазмолеммы и закрепляются в ее кортикальном слое, так что саркомеры всех миофибрилл располагаются на одном уровне. Это и создает при наблюдении в микроскоп впечатление поперечной исчерченности всего волокна.
Источником ионов кальция служат цистерны агранулярной эндоплазма-тической сети. Они вытянуты вдоль миофибрилл около каждого саркоме-ра и образуют саркоплазматическую сеть. Именно в ней аккумулируются ионы кальция, когда миосимпласт находится в расслабленном состоянии. На уровне Z-линий (у амфибии) или на границе А- и I-дисков (у млекопитающих) канальцы сети меняют направление и располагаются поперечно, образуя расширенные терминальные или латеральные (L) цистерны.
С поверхности в глубину миосимпласта плазмолемма образует длинные трубочки, идущие поперечно (Т-трубочки) на уровне границ между темными и светлыми дисками. Когда миосимпласт получает сигнал о начале сокращения, он перемещается по плазмолемме в виде потенциала действия и распространяется на мембрану Т-трубочек. Поскольку эта мембрана сближена с мембранами саркоплазматической сети, состояние последних меняется, кальций освобождается из цистерн сети и взаимодействует с актино-миозиновыми комплексами (они сокращаются). Когда потенциал действия исчезает, кальций снова аккумулируется в канальцах сети и сокращение миофибрилл прекращается. Для развития усилия сокращения нужна энергия. Она освобождается за счет превращения АТФ в АДФ. Роль АТФ-азы выполняет миозин. Источником АТФ служат главным образом митохондрии, поэтому они и располагаются непосредственно между миофибриллами.
Большую роль в деятельности миосимпластов играют включения миоглобина и гликогена. Гликоген служит источником энергии, необходимой не только для совершения мышечной работы, но и поддержания теплового баланса всего организма. Миоглобин связывает кислород, когда мышца расслаблена и через мелкие кровеносные сосуды свободно протекает кровь. Во время сокращения мышцы сосуды сдавливаются, а запасенный кислород освобождается и участвует в биохимических реакциях.
Миосателлитоциты. Эти малодифференцированные клетки являются источником регенерации мышечной ткани. Они прилежат к поверхности миосимпласта, так что их плазмолеммы соприкасаются (см. рис. 9.1, 9.4). Миосателлитоциты одноядерны, их темные ядра овальной формы и мельче, чем в симпластах. Они обладают всеми органеллами общего значения (в том числе и клеточным центром).
Типы мышечных волокон. Разные мышцы (как органы) функционируют в неодинаковых биомеханических условиях. Поэтому и мышечные волокна в составе разных мышц обладают разной силой, скоростью и длительностью сокращения, а также утомляемостью. Активность ферментов в них различна, и представлены они в различных изомерных формах. Различно в них и содержание дыхательных ферментов - гликолитических и окислительных.
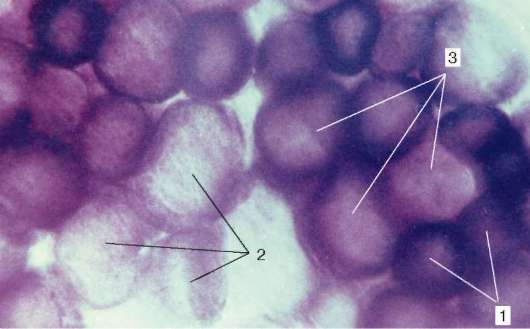
Рис. 9.7. Активность сукцинатдегидрогеназы в мышечных волокнах разного типа (препарат В. Ф. Четвергова, обработка по Нахласу и соавт.): 1 - высокая; 2 - низкая; 3 - средняя
По соотношению миофибрилл, митохондрий и миоглобина различают белые, красные и промежуточные волокна. По функциональным особенностям мышечные волокна подразделяют на быстрые, медленные и промежуточные, что определяется молекулярной организацией миозина. Среди его изоформ существуют две основные - «быстрая» и «медленная». При постановке гистохимических реакций их идентифицируют по АТФ-азной активности. С этими свойствами коррелирует и активность дыхательных ферментов. Обычно в быстрых волокнах преобладают гликолитические процессы, они богаты гликогеном, в них меньше миоглобина, поэтому их называют белыми. В медленных волокнах, напротив, выше активность окислительных ферментов, они богаче миоглобином, выглядят более красными.
Наряду с белыми и красными существуют и промежуточные волокна. В составе большинства скелетных мышц волокна разного гистохимического типа располагаются мозаично (рис. 9.7).
Свойства мышечных волокон меняются при изменении нагрузок - спортивных, профессиональных, а также в экстремальных условиях (невесомость). При возврате к обычной деятельности такие изменения обратимы. При некоторых заболеваниях (мышечные атрофии, дистрофии, последствия денервации) мышечные волокна с разными исходными свойствами изменяются неодинаково. Это позволяет уточнять диагноз, для чего исследуют биоптаты скелетных мышц.
Регенерация. Ядра миосимпластов делиться не могут, так как в саркоплазме отсутствуют клеточные центры. Камбиальными элементами служат миосателлитоциты. Пока организм растет, они делятся, а дочерние клетки сливаются с миосимпластами. По окончании роста размножение миосателлитоцитов затухает. После повреждения мышечного волокна на некотором протяжении от места травмы оно разрушается и его фрагмен-
ты фагоцитируются макрофагами. Восстановление тканей осуществляется за счет двух механизмов: компенсаторной гипертрофии самого симпласта и пролиферации миосателлитоцитов. В симпласте активизируются гранулярная эндоплазматическая сеть и комплекс Гольджи. Происходит синтез веществ, необходимых для восстановления саркоплазмы и миофибрилл, а также сборка мембран, так что восстанавливается целостность плазмо-леммы. Поврежденный конец миосимпласта при этом утолщается, образуя мышечную почку. Миосателлитоциты, сохранившиеся рядом с повреждением, делятся. Одни из них мигрируют к мышечной почке и встраиваются в нее, другие сливаются (так же, как миобласты при гистогенезе) и образуют новые миотубы, которые развиваются в мышечных волокна.
9.2.2. Скелетная мышца как орган
Передача усилий сокращения на скелет осуществляется посредством сухожилий или прикрепления мышц непосредственно к надкостнице. На конце каждого мышечного волокна плазмолемма образует глубокие узкие впячивания. В них со стороны сухожилия или надкостницы проникают тонкие коллагеновые волокна. Последние спирально оплетаются ретикулярными волокнами. Концы волокон направляются к базальной мембране, входят в нее, поворачивают назад и по выходе снова оплетают коллагеновые волокна соединительной ткани.
Между мышечными волокнами находятся тонкие прослойки рыхлой волокнистой соединительной ткани - эндомизий. Коллагеновые волокна наружного листка базальной мембраны вплетаются в него (см. рис. 9.3, в), что способствует объединению усилий при сокращении миосимпластов. Более толстые прослойки рыхлой соединительной ткани окружают по несколько мышечных волокон, образуя перимизий и разделяя мышцу на пучки. Несколько пучков объединяются в более крупные группы, разделенные более толстыми соединительнотканными прослойками. Соединительную ткань, окружающую поверхность мышцы, называют эпимизием.
Васкуляризация. Артерии вступают в мышцу и распространяются по прослойкам соединительной ткани, постепенно истончаясь. Ветви пятого-шестого порядка образуют в перимизии артериолы. В эндомизии расположены капилляры. Они идут вдоль мышечных волокон, анастомозируя друг с другом. Венулы, вены и лимфатические сосуды проходят рядом с приносящими сосудами. Как обычно, рядом с сосудами много тучных клеток, принимающих участие в регуляции проницаемости сосудистой стенки.
Иннервация. В мышцах выявлены миелинизированные эфферентные (двигательные), афферентные (чувствительные), а также немиелинизиро-ванные вегетативные нервные волокна. Отросток нервной клетки, приносящий импульс от мотонейрона спинного мозга, ветвится в перимизии. Каждая его ветвь проникает сквозь базальную мембрану и у поверхности симпласта на плазмолемме образует терминали, участвуя в организации так называемой моторной бляшки (см. главу 10, рис. 10.18). При поступлении
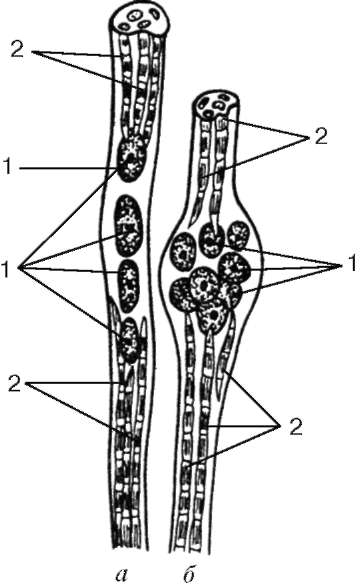
Рис. 9.8. Фрагмент мышечного веретена, содержащего мышечные волокна с ядерной цепочкой (а) и с ядерной сумкой (б) (схема по Г. С. Катинасу): 1 - ядра; 2 - миофибриллы (органеллы общего значения не показаны)
нервного импульса из терминалей выделяется ацетилхолин - медиатор, который вызывает возбуждение (потенциал действия), распространяющееся отсюда по плазмолемме миосимпласта.
Итак, каждое мышечное волокно иннервируется самостоятельно и окружено сетью гемокапилляров, образуя комплекс, именуемый мионом.
Группа мышечных волокон, ин-нервируемых одним мотонейроном, называется нервно-мышечной единицей. Мышечные волокна, принадлежащие к одной нервно-мышечной единице, лежат не рядом, а расположены мозаично среди волокон, относящихся к другим единицам.
Чувствительные нервные окончания располагаются не на рабочих (экстрафузальных) мышечных волокнах, а связаны со специализированными мышечными волокнами в так называемых мышечных вере-
тенах (с интрафузальными мышечными волокнами), которые расположены в перимизии.
Интрафузальные мышечные волокна. Интрафузальные мышечные волокна веретен значительно тоньше рабочих. Существует два их вида - волокна с ядерной сумкой и волокна с ядерной цепочкой (рис. 9.8). Ядра в тех и в других округлые и расположены в толще симпласта, а не у его поверхности. В волокнах с ядерной сумкой ядра симпласта образуют скопления в его утолщенной средней части. В волокнах с ядерной цепочкой в средней части симпласта утолщение не образуется, ядра лежат здесь продольно одно за другим. Рядом со скоплениями ядер расположены органеллы общего значения.
Миофибриллы находятся в концах симпластов. Сарколемма волокна соединяется с капсулой нервно-мышечного веретена, состоящей из плотной волокнистой соединительной ткани. Каждое мышечное волокно веретена спирально обвито терминалью чувствительного нервного волокна. В результате сокращения или расслабления рабочих мышечных волокон изменяется натяжение соединительнотканной капсулы веретена, соответственно изменяется тонус интрафузальных мышечных волокон. Вследствие этого возбуждаются чувствительные нервные окончания, обвивающие их, и в области терминалей возникают афферентные нервные импульсы. На каждом миосимпласте располагается также своя моторная бляшка. Именно поэтому интрафузальные мышечные волокна постоянно находятся в напряжении, подстраиваясь к длине мышечного брюшка в целом.
9.2.3. Сердечная мышечная ткань
Гистогенез и виды клеток. Источники развития сердечной поперечнополосатой мышечной ткани (textus muscularis striatus cardiacus) - симметричные участки висцерального листка спланхнотома в шейной части зародыша - миоэпикардиальные пластинки. Из них дифференцируются также клетки мезо-телия эпикарда. Исходные клетки сердечной мышечной ткани - кардиомио-бласты - характеризуются рядом признаков: клетки уплощены, содержат крупное ядро, светлую цитоплазму, бедную рибосомами и митохондриями. В дальнейшем происходит развитие комплекса Гольджи, гранулярной эндо-плазматической сети. В кардиомиобластах обнаруживаются фибриллярные структуры, но миофибрилл нет. Клетки обладают высоким пролифератив-ным потенциалом.
После ряда митотических циклов кардиомиобласты дифференцируются в кардиомиоциты, в которых начинается саркомерогенез (рис. 9.9). В цитоплазме кардиомиоцитов увеличивается число полисом, канальцев гранулярной эндоплазматической сети, накапливаются гранулы гликогена, возрастает объем актомиозинового комплекса. Кардиомиоциты сокращаются, но не теряют способность к дальнейшей пролиферации и дифференцировке. Развитие сократительного аппарата в позднем эмбриональном и постна-тальном периодах происходит путем надставки новых саркомеров и наслоения вновь синтезированных миофиламентов.
Дифференцировка кардиомиоцитов сопровождается увеличением числа митохондрий, распределением их у полюсов ядер и между миофи-бриллами и протекает параллельно со специализацией контактирующих поверхностей клеток. Кардиомиоциты путем контактов «конец в конец», «конец в бок» формируют сердечные мышечные волокна, и в целом ткань представляет собой сетевидную структуру. Часть кардиомиоцитов на ранних этапах кардиомиогенеза являются сократительно-секреторными. В дальнейшем в результате дивергентной дифференцировки возникают «темные» (сократительные) и «светлые» (проводящие) миоциты, в которых исчезают секреторные гранулы, тогда как в предсердных мио-цитах они сохраняются. Так формируется дифферон эндокринных кардиомиоцитов. Эти клетки содержат центрально расположенное ядро с диспергированным хроматином, одним-двумя ядрышками. В цитоплазме хорошо развиты гранулярная эндоплазматическая сеть, диктиосомы комплекса Гольджи, в тесной связи с элементами которого находятся многочисленные секреторные гранулы диаметром около 2 мкм, содержащие электронно-плотный материал. В дальнейшем секреторные гранулы обнаруживаются под сарколеммой и выделяются в межклеточное пространство путем экзоцитоза.
В целом в ходе гистогенеза возникает пять видов кардиомиоцитов - рабочие (сократительные), синусные (пейсмекерные), переходные, проводящие, а также секреторные. Рабочие (сократительные) кардиомиоциты образуют свои цепочки (рис. 9.10). Именно они, укорачиваясь, обеспечивают силу сокращения всей сердечной мышцы. Рабочие кардиомиоциты способны
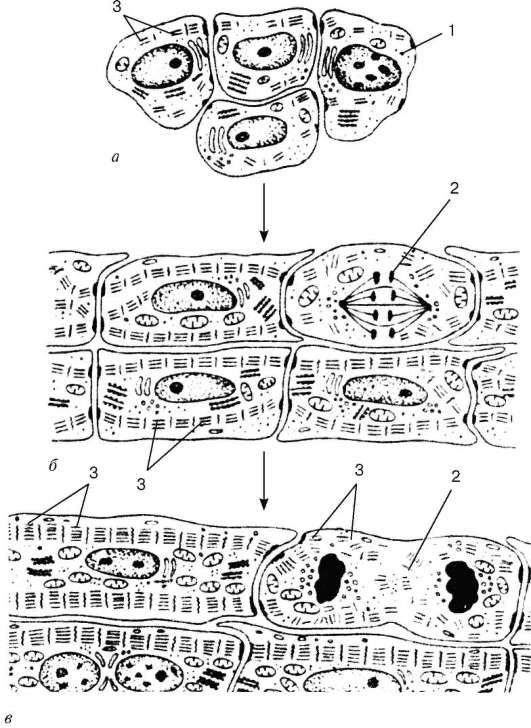
Рис. 9.9. Гистогенез сердечной мышечной ткани (схема по П. П. Румянцеву): а - кардиомиоциты в стенке сердечной трубки; б - кардиомиоциты в позднем эмбриогенезе; в - кардиомиоциты в постанатальном периоде. 1 - кардиомиоцит; 2 - митотически делящийся кардиомиоцит; 3 - миофиламенты и миофибриллы
передавать управляющие сигналы друг другу. Синусные (пейсмекерные) кардиомиоциты способны автоматически в определенном ритме сменять состояние сокращения на состояние расслабления. Клетки воспринимают управляющие сигналы от нервных волокон, в ответ на что изменяют ритм сократительной деятельности. Синусные (пейсмекерные) кардиомиоциты передают управляющие сигналы переходным кардиомиоцитам, а последние - проводящим и рабочим кардиомиоцитам. Проводящие кардиомиоциты образуют цепочки клеток, соединенных своими концами, и располагаются под эндо-
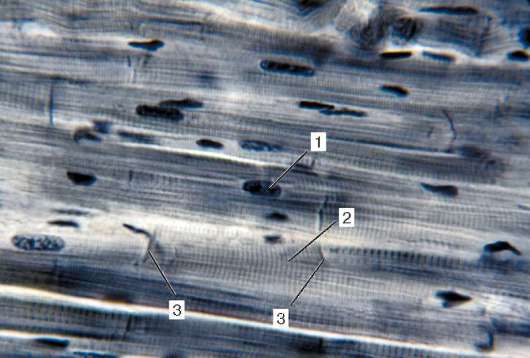
Рис. 9.10. Строение сердечной мышечной ткани (микрофотография). Окраска - железный гематоксилин:
1 - ядро кардиомиоцита; 2 - цепочка кардиомиоцитов; 3 - вставочные диски
кардом. Первая клетка в цепочке воспринимает управляющие сигналы от синусных кардиомиоцитов и передает их далее - другим проводящим кар-диомиоцитам. Клетки, замыкающие цепочку, передают сигнал через переходные кардиомиоциты рабочим. Секреторные кардиомиоциты выполняют особую функцию. Они вырабатывают пептидный гормон кардиодилатин, который циркулирует в крови в виде кардионатрина, вызывает сокращение гладких миоцитов артериол, увеличение почечного кровотока, ускоряет клубочковую фильтрацию и выделение натрия. Все кардиомиоциты покрыты базальной мембраной.
Строение сократительных (рабочих) кардиомиоцитов. Клетки имеют удлиненную (100-150 мкм) форму, близкую к цилиндрической. Их концы соединяются друг с другом, так что цепочки клеток составляют так называемые функциональные волокна (толщиной до 20 мкм). В области контактов клеток образуются так называемые вставочные диски (рис. 9.10). Кардиомиоциты могут ветвиться и образуют пространственную сеть. Их поверхности покрыты базальной мембраной, в которую снаружи вплетаются ретикулярные и коллагеновые волокна. Ядро кардиомиоцита (иногда их два) овальное и лежит в центральной части клетки. У полюсов ядра сосредоточены немногочисленные органеллы общего значения, за исключением агранулярной эндоплазматической сети и митохондрий.
Специальные органеллы, которые обеспечивают сокращение, называются миофибриллами. Они слабо обособлены друг от друга, могут расщепляться. Их строение аналогично строению миофибрилл миосимпласта скелетного мышечного волокна. Каждая митохондрия располагается на протяжении всего саркомера. От поверхности плазмолеммы в глубь кардиомиоцита направлены Т-трубочки, находящиеся на уровне Z-линии. Их мембраны сближены,
контактируют с мембранами гладкой эндоплазматической (саркоплазмати-ческой) сети. Петли последней вытянуты вдоль поверхности миофибрилл и имеют латеральные расширения (L-системы), формирующие вместе с Т-трубочками триады или диады (рис. 9.11, а). В цитоплазме имеются включения гликогена и липидов, особенно много включений миоглобина. Механизм сокращения кардиомиоцитов такой же, как у миосимпласта.
Организация кардиомиоцитов в ткань. Кардиомиоциты соединяются друг с другом по типу «конец в конец». Здесь образуются вставочные диски: эти участки выглядят как тонкие пластинки при среднем увеличении светового микроскопа. Фактически же концы кардиомиоцитов имеют неровную поверхность, поэтому выступы одной клетки входят во впадины другой. Поперечные участки выступов соседних клеток соединены друг с другом интердигитациями и десмо-сомами (рис. 9.11, б).
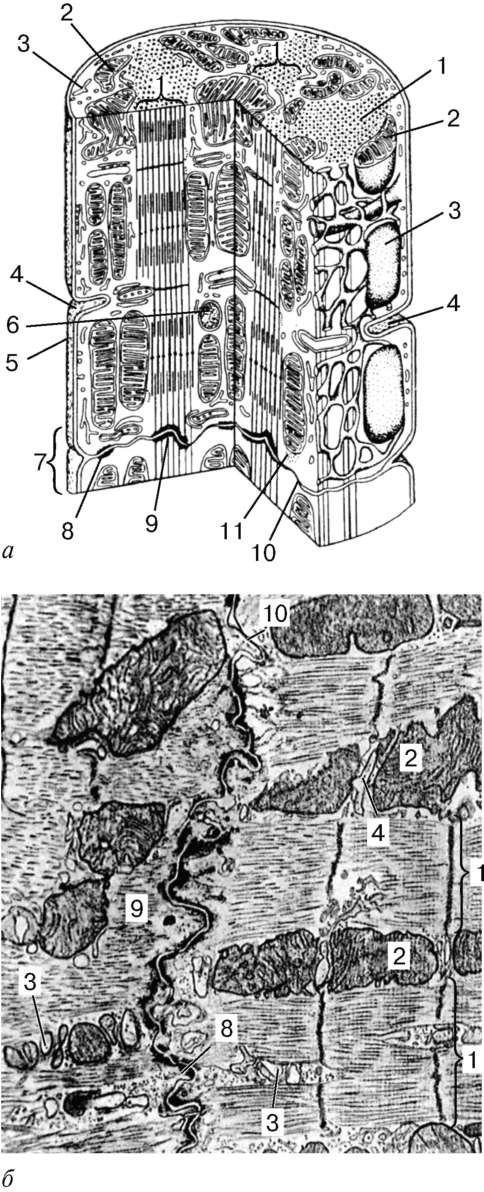
Рис. 9.11. Строение кардиомиоцита: а - схема (по Ю. И. Афанасьеву и В. Л. Горячкиной); б - электронная микрофотография вставочного диска. Увеличение 20 000. 1 - миофибриллы; 2 - митохондрии; 3 - саркотубулярная сеть; 4 - Т-трубочки; 5 - базальная мембрана; 6 - лизосома; 7 - вставочный диск; 8 - десмосома; 9 - зона прикрепления миофибрилл; 10 - щелевые контакты; 11 - гликоген
К каждой десмосоме со стороны цитоплазмы подходит миофибрил-ла, закрепляющаяся концом в десмо-плакиновом комплексе. Таким образом, при сокращении тяга одного кардиомиоцита передается другому. Боковые поверхности выступов кардиомиоцитов объединяются нексусами (щелевыми соединениями). Это создает между ними метаболические связи и обеспечивает синхронность сокращений.
Регенерация. В гистогенезе сердечной мышечной ткани камбий не возникает. Поэтому регенерация ткани протекает на основе внутриклеточных гиперпластических процессов. Вместе с тем для кардиомиоцитов млекопитающих, приматов и человека характерен процесс полиплоиди-
зации. Например, у обезьян ядра до 50 % терминально дифференцированных кардиомиоцитов становятся тетра- и октоплоидными. Полиплоидные кардиомиоциты возникают за счет ацитокинетического митоза, что приводит к многоядерности. В условиях патологии сердечно-сосудистой системы человека (ревматизм, врожденные пороки сердца, инфаркт миокарда и др.) важную роль в компенсации повреждений кардиомиоцитов играют внутриклеточная регенерация, полиплоидизация ядер, возникновение многоядерных кардиомиоцитов.
9.3. ГЛАДКИЕ МЫШЕЧНЫЕ ТКАНИ
Различают три группы гладких (неисчерченных) мышечных тканей (textus muscularis nonstriatus) и клеток: мезенхимные, нейральные и миоэпи-телиальные клетки.
9.3.1. Мышечная ткань мезенхимного происхождения
Гистогенез. Эта ткань делится на два вида: висцеральную и сосудистую. В эмбриональном гистогенезе даже электронно-микроскопически трудно отличить мезенхимные предшественники фибробластов от гладких миоци-тов. В малодифференцированных гладких миоцитах развиты гранулярная эндоплазматическая сеть, комплекс Гольджи. Тонкие филаменты ориентированы вдоль длинной оси клетки. По мере развития размеры клетки и число филаментов в цитоплазме возрастают. Постепенно объем цитоплазмы, занятый сократительными филаментами, увеличивается, расположение их в цитоплазме становится все более упорядоченным. Пролиферативная активность гладких миоцитов в миогенезе постепенно снижается. Это происходит в результате увеличения продолжительности клеточного цикла, выхода клеток из цикла репродукции и перехода в дифференцированное состояние. Дифференцируясь, они синтезируют компоненты межклеточного матрикса, коллаген базальной мембраны, а также эластин. У дефинитивных клеток (миоцитов) синтетическая способность снижена, но не исчезает полностью.
Строение и функционирование клеток. Гладкий миоцит - веретеновидная клетка длиной 20-500 мкм, шириной 5-8 мкм. Ядро палочковидное, находится в ее центральной части. Когда миоцит сокращается, его ядро изгибается и даже закручивается (рис. 9.12-9.14).
Структура дефинитивных гладких миоцитов (лейомиоцитов), входящих в состав внутренних органов и стенки сосудов, имеет много общего, но в то же время характеризуется гетероморфией. Так, в стенках вен и артерии обнаруживаются овоидные, веретеновидные, отростчатые миоциты длиной 10-40 мкм, доходящие иногда до 140 мкм. Наибольшей длины гладкие мио-
циты достигают в стенке матки - до 500 мкм. Диаметр миоцитов колеблется от 2 до 20 мкм. В зависимости от характера внутриклеточных биосинтетических процессов различают контрактильные и секреторные миоциты. Первые специализированы на функции сокращения, но вместе с тем сохраняют секреторную активность.
Секреторные миоциты по своей ультраструктуре напоминают фиб-робласты, однако содержат в своей цитоплазме пучки тонких миофи-ламентов, расположенные на периферии клетки. В цитоплазме хорошо развиты комплекс Гольджи, гранулярная эндоплазматическая сеть, много митохондрий, гранул гликогена, свободных рибосом и полисом. По степени зрелости такие клетки относят к малодифференцирован-ным. Филаменты актина образуют в цитоплазме трехмерную сеть, вытянутую преимущественно продольно. Концы филаментов скреплены между собой и с плазмолеммой специальными сшивающими белками. Эти участки хорошо видны на электронных микрофотографиях как плотные тельца. Мономеры миозина располагаются рядом с филаментами актина. Плазмолемма образует впя-чивания - кавеолы, в которых концентрируются ионы кальция. Сигнал к сокращению обычно поступает по нервным волокнам. Медиатор, который выделяется из их термина-лей, изменяет проницаемость плаз-молеммы. Происходит высвобождение ионов кальция, что влечет за собой как полимеризацию миозина, так и взаимодействие миозина с актином.
Происходит втягивание актино-вых миофиламентов между миози-
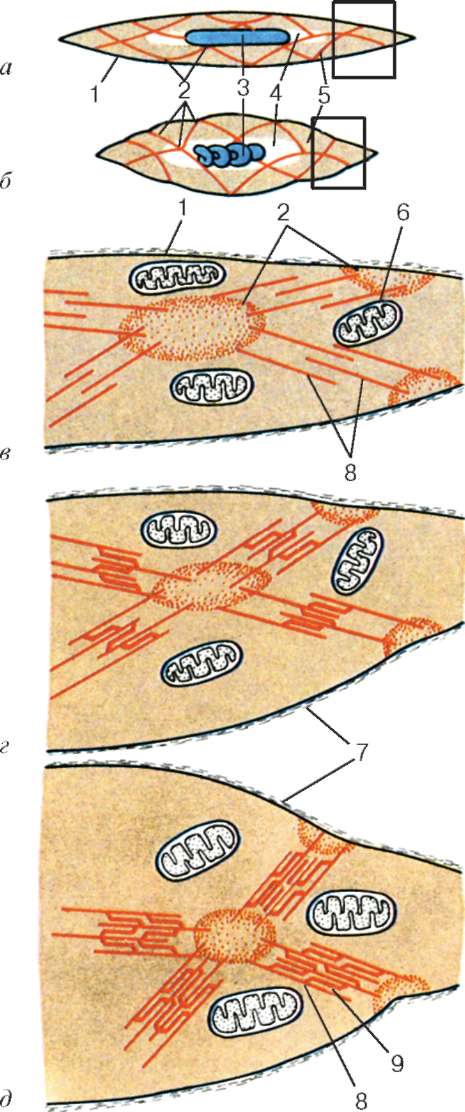
Рис. 9.12. Строение гладкого миоцита (схема):
а, в - при расслаблении; б, д - при наибольшем сокращении; г - при неполном сокращении; в-д - увеличенные изображения участков, обведенных рамками на фрагментах а и б. 1 - плазмолемма; 2 - плотные тельца; 3 - ядро; 4 - эндоплазма; 5 - сократительные комплексы; 6 - митохондрии; 7 - базальная мембрана; 8 - актиновые (тонкие) мио-филаменты; 9 - миозиновые (толстые) миофиламенты
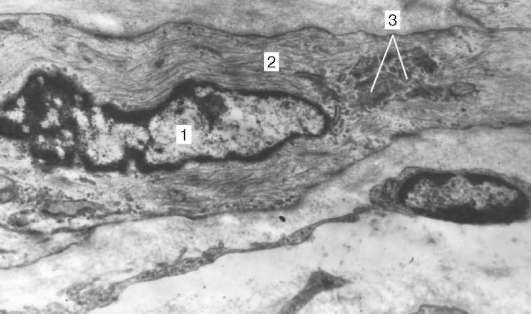
Рис. 9.13. Ультраструктура дифференцирующегося гладкого миоцита в стенке бронха:
1 - ядро; 2 - цитоплазма с миофиламентами; 3 - комплекс Гольджи, увеличение 35 000 (препарат А. Л. Зашихина)
новыми, плотные пятна сближаются, усилие передается на плазмолемму, и вся клетка укорачивается (см. рис. 9.12). Когда поступление сигналов со стороны нервной системы прекращается, ионы кальция перемещаются из цитоплазмы в кавеолы и в канальцы эндоплазматической сети, миозин деполимеризуется и «миофибриллы» распадаются. Сокращение прекращается. Таким образом, актиномиозиновые комплексы существуют в гладких миоцитах только в период сокращения при наличии в цитоплазме свободных ионов кальция.
Миоциты окружены базальной мембраной. На отдельных участках в ней образуются «окна», поэтому плазмолеммы соседних миоцитов сближаются. Здесь формируются нексусы, и между клетками возникают не только механические, но и метаболические связи. Поверх «чехликов» из базальной мембраны между миоцитами проходят эластические и ретикулярные волокна, объединяющие клетки в единый тканевой комплекс. Гладкие миоциты синтезируют протеогликаны, гликопротеиды, проколлаген, проэластин, из которых формируются коллагеновые и эластические волокна и аморфный компонент межклеточного матрикса. Взаимодействие миоцитов осуществляется при помощи цитоплазматических мостиков, взаимных впячиваний, нексусов, десмосом, участков мембранных контактов поверхностей миоцитов.
Регенерация. Гладкая мышечная ткань висцерального и сосудистого видов обладает значительной чувствительностью к воздействию экстремальных факторов. В активированных миоцитах возрастает уровень биосинтетических процессов, морфологическим выражением которых являются синтез сократительных белков, укрупнение и гиперхроматоз ядра, гипертрофия ядрышка, возрастание показателей ядерно-цитоплазматического отношения, увеличение количества свободных рибосом и полисом, актива-

Рис. 9.14. Строение гладкой мышечной ткани (объемная схема) (по Р. В. Крстичу, с изменениями):
1 - веретеновидные гладкие миоциты; 2 - цитоплазма миоцита; 3 - ядра миоци-тов; 4 - плазмолемма; 5 - базальная мембрана; 6 - поверхностные пиноцитозные пузырьки; 7 - межклеточные соединения; 8 - нервное окончание; 9 - коллагеновые фибриллы; 10 - микрофиламенты
ция ферментов, аэробного и анаэробного фосфорилирования, мембранного транспорта. Клеточная регенерация осуществляется как за счет дифференцированных клеток, обладающих способностью вступать в митотический цикл, так и за счет активизации камбиальных элементов (миоцитов малого объема). При действии ряда повреждающих факторов отмечается феноти-пическая трансформация контрактильных миоцитов в секреторные. Данная трансформация часто наблюдается при повреждении интимы сосудов, формировании интимальной гиперплазии при развитии атеросклероза.
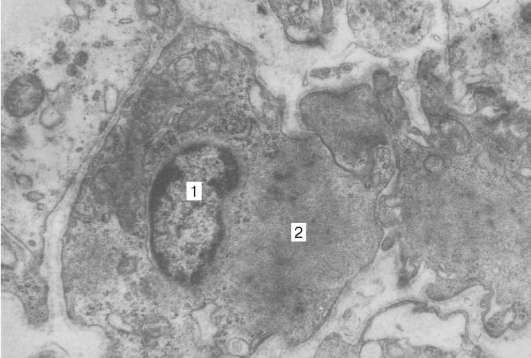
Рис. 9.15. Ультраструктура миопигментоцита (препарат Н. Н. Сарбаевой): 1 - ядро; 2 - миофиламенты, увеличение 6000
9.3.2. Мышечная ткань мезенхимного типа в составе органов
Миоциты объединяются в пучки, между которыми располагаются тонкие прослойки соединительной ткани. В эти прослойки вплетаются ретикулярные и эластические волокна, окружающие миоциты. В прослойках проходят кровеносные сосуды и нервные волокна. Терминали последних оканчиваются не непосредственно на миоцитах, а между ними. Поэтому после поступления нервного импульса медиатор распространяется диффуз-но, возбуждая сразу многие клетки. Гладкая мышечная ткань мезенхимно-го происхождения представлена главным образом в стенках кровеносных сосудов и многих полых внутренних органов.
Гладкая мышечная ткань в составе конкретных органов имеет неодинаковые функциональные свойства. Это обусловлено тем, что на поверхности органов имеются разные рецепторы конкретных биологически активных веществ. Поэтому и на многие лекарственные препараты их реакция неодинакова. Возможно, разные функциональные свойства тканей связаны и с конкретной молекулярной организацией актиновых филаментов.
9.3.3. Мышечная ткань нейрального происхождения
Мышечная ткань радужки и цилиарного тела относится к четвертому типу сократимых тканей. Миоциты этой ткани развиваются из клеток нейрального зачатка в составе внутренней стенки глазного бокала. В ряду
Рис. 9.16. Миоэпителиальные клетки в концевом отделе слюнной железы (схема по Г. С. Катинасу):
а - поперечный срез; б - вид с поверхности. 1 - ядра миоэпителиоцитов; 2 - отростки миоэпителиоцитов; 3 - ядра секреторных эпителиоцитов; 4 - базальная мембрана
позвоночных мышечные элементы радужки обнаруживают разнообразную дивергентную дифференцировку. Так, мионейральная ткань у рептилий и птиц представлена исчерченными многоядерными волокнами, имеющими большое сходство с мускулатурой скелетного типа. У млекопитающих и человека основной структурно-функциональной единицей мышц радужки является гладкий одноядерный миоцит, или миопигментоцит. Последние имеют пигментированное тело, содержащее одно ядро, вынесенное за пределы веретеновидной сократимой части (рис. 9.15).
Цитоплазма клеток содержит большое число митохондрий и пигментные гранулы, которые сходны по размерам и форме с гранулами пигментного эпителия. Миофиламенты в миопигментоцитах делятся на тонкие (7 нм) и толстые (1,5 нм), по размерам и расположению напоминают миофиламен-ты гладких миоцитов. Каждый миопигментоцит окружен базальной мембраной. Возле цитоплазматических отростков миоцитов обнаруживаются безмиелиновые нервные волокна. В зависимости от направления отростков (перпендикулярно или параллельно краю зрачка) миоциты образуют две мышцы - суживающую и расширяющую зрачок.
Регенерация. В немногочисленных работах показана низкая регенераци-онная активность после повреждения или ее отсутствие.
9.3.4. Мышечные клетки эпидермального происхождения
Миоэпителиальные клетки развиваются из эпидермального зачатка. Они встречаются в потовых, молочных, слюнных и слезных железах и имеют общих предшественников с их секреторными клетками. Миоэпителиальные
клетки непосредственно прилежат к собственно эпителиальным и имеют общую с ними базальную мембрану. При регенерации те и другие клетки восстанавливаются тоже из общих малодифференцированных предшественников. Большинство миоэпителиальных клеток имеют звездчатую форму. Эти клетки нередко называют корзинчатыми: их отростки охватывают концевые отделы и мелкие протоки желез (рис. 9.16). В теле клетки располагаются ядро и органеллы общего значения, а в отростках - сократительный аппарат, организованный как и в клетках мышечной ткани мезенхимного типа.
Контрольные вопросы
1. Генетическая классификация мышечных тканей. Структурно-функциональные единицы разных типов мышечной ткани.
2. Поперечнополосатая скелетная мышечная ткань: развитие, строение, морфологические основы мышечного сокращения. Регенерация.
3. Поперечнополосатая сердечная мышечная ткань: развитие, специфика строения различных видов кардиомиоцитов, регенерация.
4. Разновидности гладких миоцитов: источники развития, топография в организме, регенерация.
Гистология, эмбриология, цитология: учебник / Ю. И. Афанасьев, Н. А. Юрина, Е. Ф. Котовский и др.. - 6-е изд., перераб. и доп. - 2012. - 800 с. : ил.
Мышцы тела человека образованы в основном мышечной тканью, состоящей из мышечных клеток. Различают гладкую и поперечнополосатую мышечную ткань. Гладкая мышечная ткань бразует мускулатуру, которая входит в состав некоторых внутренних органов, а поперечнополосатая образует скелетные мышцы. Общим свойством мышечной ткани является её возбудимость , проводимость и сократимость .
Поперечнополосатая мышечная ткань отличается от гладкой более высокой возбудимостью, проводимостью и сократимостью. Клетки поперечнополосатой мускулатуры имеют очень малый диаметр и большую длину (до 10-12 см). В связи с этим их называют волокнами .
Как и другие клетки, мышечные клетки имеют протоплазму, которая называется саркоплазмой (от греч. саркос - мясо). Мембрана мышечных клеток называется сарколеммой . Внутри мышечного волокна находятся многочисленные ядра и другие составные части клеток.
В состав мышечных волокон входит большое количество еще более тонких волоконец - миофибрилл , которые, в свою очередь, состоят из тончайших нитей - протофибрилл. Протофибриллы - это сократительный аппарат мышечной клетки, они представляют собой специальные сократительные белки - миозин и актин . Механизм мышечных сокращений представляет собойсложный процесс физических и химических превращений в мышечном волокне при обязательном участии сократительного аппарата. Запуск этого механизма осуществляется нервным импульсом, а энергия для процесса сокращения поставляется аденозинтрифосфорной кислотой(АТФ). В этой связи особенностью строения мышечных волокон является также большой количество митохондрий, обеспечивающих мышечное волокно необходимой энергией. Расслабление мышечного волокна, по предположению многих ученых, осуществляется пассивно, благодаря эластичности сарколеммы и внутримышечной соединительной ткани.
Строение, форма и классификация скелетных мышц
Анатомической единицей самой активной части мышечной системы человека - скелетной, или поперечнополосатой, мускулатуры - является скелетная мышца. Скелетная мышца - это орган, образованный поперечнополосатой мышечной тканью, содержащей, кроме того, соединительную ткань, нервы и сосуды.
Каждая мышца окружена своеобразным "футляром" из соединительной ткани - фасцией .
Во внешнем строении мышцы различают сухожильную головку, соответствующую началу мышцы, брюшко мышцы, или тело, образованное мышечными волокнами, и сухожильный конец мышцы, или хвост, с помощью которого мышца прикрепляется к другой кости. Обычно хвост мышцы является подвижной точкой прикрепления, а начало - неподвижной. В процессе движения их функции могут меняться: подвижные точки становятся неподвижными и наоборот.
Форма мышц очень разнообразна и в значительной степени зависит от функционального назначения мышцы. Различают длинные, короткие, широкие, круговые и другие мышцы. Длинные мышцы располагаются на конечностях, короткие - там, гдеразмах движения мал (например, между позвонками). Широкие мышцы располагаются преимущественно на туловище (мышцы живота, спины, груди). Круговые мышцы располагаются вокруг отверстий тела и при сокращении суживают их. Такие мышцы называют сфинктерами . Если мышца имеет одну головку, её называют простой, если две или больше - сложной (например, двуглавая, трёхглавая и четырёхглавая мышцы). Важным морфологическим признаком является расположение мышечных волокон. Различают параллельное, косое, поперечное и круговое расположение волокон (у сфинктеров). если при косом расположении мышечных волокон они присоединяются только с одной стороны сухожилиями, то мышцы называют одноперистыми , если с двух сторон - двуперистыми .
Функционально мышцы можно разделить на сгибатели и разгибатели , вращатели снаружи и вращатели внутри , приводящие мышцы и отводящие . Выделяют также мышцы - синергисты и мышцы - антагонисты . Первые образуют группу мышц, содружественно выполняющих какое-либо движение, сокращение вторых вызывает противоположные движения.
Основные группы мышц
Скелетные мышцы взрослого человека составляют около 40% массы его тела. У новорождённых и детей раннего возраста мышцы составляют не более 20-25% массы тела, а в пожилом возрасте отмечается постепенное уменьшение массы мускулатуры до 25-30% от массы тела. Всего в теле человека насчитывается более 600 мышц.
По месту расположения, т.е. по топографическому признаку, выделяют мышцы спины, груди, живота, головы, шеи, верхних и нижних конечностей.
Мышцы спины делятся на поверхностные и глубокие . К поверхностным мышцам спины относятся: трапециевидная мышца, широчайшая мышца спины, ромбовидные мышцы (большая и малая), зубчатые мышцы (верхняя и нижняя).
Эти мышцы участвуют в движениях верхних конечностей и изменении объёма грудной клетки (акт дыхания).
Глубокие мышцы спины представлены многочисленными мышцами, расположенными вдоль позвоночного столба. Они прикрепляются главным образом к отросткам позвонков и участвуют в движениях позвоночника назад и в сторону.
Мышцы груди делятся на мышцы, относящиеся к плечевому поясу и верхней конечности (большая и малая грудные мышцы, подключичная и передняя зубчатая мышцы), и собственные мышцы груди (наружные внутренние межрёберные мышцы). Они приводят в движение и укрепляют на туловище верхние конечности и участвуют в функции дыхания. К дыхательным мышцам принадлежит и диафрагма (она отделяет грудную полость от брюшной).
Мышцы живота представлены наружной и внутренней косыми, поперечной и прямой мышцами живота, а также квадратной мышцей поясницы.
Прямые мышцы живота участвуют в сгибании туловища вперёд, а косые мышцы обеспечивают наклоны позвоночника в стороны и его повороты вместе с грудной клеткой вправо и влево. Мышцы живота не только участвуют в движениях туловища и грудной клетки. Они образуют стенку брюшной полости и, благодаря своему тонусу, удерживают внутренние органыв их нормальном положении. Своим сокращением они повышают внутрибрюшное давление, образуя так называемый брюшной пресс . Мышцы брюшного пресса облегчают опорожнение кишечника (дефекацию), мочеиспускание, а у женщин изгнание плода при родах. Сокращение мышц брюшного пресса также способствует движению крови в венозной системе и осуществлению дыхательных движений, но и образование грыжи. При грыже происходит выпячивание внутренних органов (кишечника, желудка и др.) из брюшной полости под кожу.
Среди мышц головы различают жевательные мышцы и мимические. К жевательным мышцам относится височная, жевательная, крыловидные. Сокращение этих мышц вызывает сложные жевательные движения нижней челюсти. Мимические мышцы отличаются от других мышц человеческого тела тем, что одним, а иногда и двумя своими концами прикрепляются к коже лица. Сокращение этих мышц смещение кожи, образование складок и морщин. Это определяет мимику, т.е. то или иное выражение лица. Проявление сложных ощущений (эмоций) - радости, стыда, презрения, горя и боли и т.д. - определяется многочисленными комбинациями сокращений мимических мышц, подчиненных импульсам, идущим к ним от коры головного мозга по лицевому нерву. Круговые мышцы глаза и рта также относятся к числу мимических мышц. Мышцы шеи запрокидывают голову, наклоняют её и поворачивают. Мышцы, прикрепленные к подъязычной кости, при сокращении изменяют положение языка и гортани при глотании и произнесении звуков.
Мышцы верхней конечности подразделяются на мышцы плечевого пояса и мышцы свободной верхней конечности. Мышцы плечевого пояса окружают плечевой сустав, обеспечивая многочисленные движения в нём. Среди них важнейшая - дельтовидная мышца. При сокращении эта мышца сгибает руку плечевом суставе и отводит руку до горизонтального положения.
К мышцам свободной верхней конечности относятся мышцы плеча, предплечья и кисти.
В области плеча спереди расположена группа мышц - сгибателей , сзади - разгибателей . Среди мышц передней группы наиболее важной является двуглавая мышца плеча (бицепс), задней - трёхглавая мышца плеча.
Мышцы предплечья также на передней поверхности представлены сгибателями, на задней - разгибателями. Мышцы кисти расположены только на ладонной поверхности. Среди них выделяются - длинная ладонная мышца и сгибатели пальцев. Благодаря наличию собственного мышечного аппарата, пальцы кисти, особенно большой палец, приобретают большую подвижность и способны к разнообразным движениям, в том числе к максимальному сгибанию и разгибанию, что чрезвычайно важно при работе.
Мышцы, расположенные в области пояса нижних конечностей, приводят в движение ногу в тазобедренном суставе, а также при фиксированной конечности сгибают позвоночник в поясничном отделе. В переднюю группу мышц входит одна крупная мышца - подвздошно-поясничная. Среди задненаружной группы мышц тазового пояса - большая, средняя и малая ягодичные мышцы.
Нижние конечности имеют более массивные скелет, чем верхние; их мускулатура обладает большой силой, но вместе с тем меньшим разнообразием и ограниченным размахом движений.
На бедре спереди располагается самая длинная в человеческом теле (до 50 см) портняжная мышца. Она сгибает ногу в тазобедренном и коленном суставах.
Четырёхглавая мышца бедра лежит глубже портняжной мышцы, облегая бедренную кость почти со всех сторон.
Основная функция этой мышцы - разгибание коленного сустава. При стоянии четырёхглавая мышца не даёт коленному суставу сгибаться.
На стопе различают тыльную и подошвенные мышцы. Эти мышцы участвуют в сгибании и разгибании пальцев стопы, а также в их сближении и разведении.