Строение черепа животных. Строение головного мозга животных и мозговые оболочки. Артерии головного мозга
СКЕЛЕТ ГОЛОВЫ. ЧЕРЕП
Появление головы и ее скелета было связано с усложнением общей организации предков позвоночных животных, происходящим в борьбе за существование и благодаря прогрессивному развитию организма, приспосабливающегося к изменяющимся условиям жизни. На переднем конце биполярного тела животного развиваются два самостоятельных раздела скелета головы (рис. 32). Один - защитное приспособление для развивающегося головного мозга - череп - cranium, второй (лицевой отдел) - в связи с совершенствованием приспособлений для лучшего, большего захвата пищи и ее первичной обработки. Так на переднем конце тела низших позвоночных возникли две части скелета головы: череп, образующий черепную полость для мозга, и висцеральный отдел, формирующий в дальнейшем лицо - fades. Эти два отдела объединили общность их расположения и тот факт, что подвижный челюстной аппарат лицевого отдела нашел опору для своих мышц на черепе.
Мозг дельфина является вторым самым мощным и сложным мозгом у животных. Интеллект может быть определен как мера способности мозга обрабатывать информацию способами, которые решают проблемы и повышают выживаемость. Измерение интеллекта может быть совершенно иным для животных, которые эволюционировали в воде по сравнению с теми, кто живет на суше, поскольку проблемы, необходимые для выживания в воде, сильно отличаются от тех, которые требуются на суше. Много дебатов существует о «интеллекте дельфинов».
Крупные мозговые животные, такие как люди, шимпанзе и дельфины, много общего, они обычно живут долгой жизнью, живут в жидких социальных группах, и в детстве они демонстрируют полную родительскую зависимость. Таким образом, невозможно полностью оценить уровень интеллекта этого животного. Но другой способ, который был предложен для измерения интеллекта, заключается в определении меры относительного размера мозга, определяемой как соотношение между фактической массой мозга и прогнозируемой массой мозга для животное данного размера.
Рис. 32. Череп акулы
Каждый отдел развивался из различных элементов, расположенных в этой области: хрящей, образующихся по бокам от хорды и впереди нее (череп), и из первых висцеральных дуг (лицо). Кости, произошедшие из этих участков, в онтогенезе проходят три стадии окостенения и являются вторичными. В дальнейшем в формировании черепа участвуют отдельные части наружного скелета, формирующие покровные кости черепа. Покровные кости черепа - первичные и в онтогенезе проходят только две стадии окостенения - перепончатую и костную (рис. 33).
Это измерение предполагает, что чем выше число, тем выше интеллект. Кроме того, степень свертывания коры головного мозга представляется мерой интеллекта. Чем больше сложенная кора, тем больше места в мозге, чтобы разместить дополнительные нейроны, с которыми можно выполнять обработку информации. Недавно опубликованная информация об увеличении складчатости коры головного мозга Альберта Эйнштейна по сравнению с другими людьми поддерживает эту теорию. Единственным животным, имеющим более сложную кору, чем человек, является дельфин.
Развитие среднего мозга
Эта картина относится к мозгу человека. основной причиной такого большого мозга является обработка информации из сложной эхолокации дельфина или биосонарной системы. Но нет доказательств, подтверждающих эту веру. Наиболее распространенная теория состоит в том, что этот более крупный мозг эволюционировал для поддержки более сложных познавательных способностей. Они могут помнить события и изучать концепции, меняя свое поведение в результате предыдущего опыта. Они могут общаться друг с другом во время кооперативного поведения, управлять отношениями в своих стручках и воспитывать своих детей.
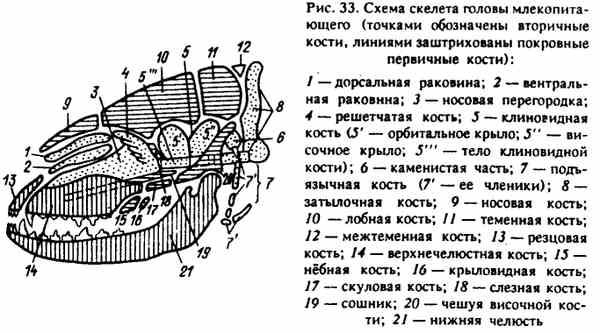
Рис. 33. Схема скелета головы млекопитающего (точками обозначены вторичные кости, линиями заштрихованы покровные первичные кости)
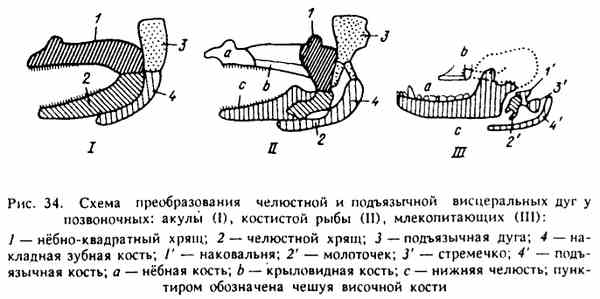
Рис. 34. Схема преобразования челюстной и подъязычной висцеральных дуг у позвоночных
Они могут понимать не только слова символического языка, но и интерпретировать синтаксический порядок языка. Это понимание синтаксиса является очень показательным для интеллекта. Подвижные свистки, создаваемые дельфинами, служат доказательством того, что дельфины имеют самосознание или способность иметь понятие я и знать, что он существует как индивидуальное существо. Самосознание существует в передней лобной коре головного мозга. Помимо дельфинов, самосознание, по-видимому, существует только у крупных мозговых приматов и человека.
В результате изучения болезни Альцгеймера у человека было обнаружено, что человеческие веретеновые нейроны ассоциируются с умением распознавать, запоминать, рассуждать, общаться, воспринимать, адаптироваться к изменениям, решать проблемы и понимать. Вспышки умершего Альцгеймера пациенты демонстрируют высокую частоту изношенных нейронов веретена. Недавно шпиндельные нейроны были изолированы в мозгу настоящих китов. Этот вывод показывает, что истинные киты обладают этими передовыми способностями, которые в прошлом были связаны только с человеком и приматами.
Череп формировался из множества костей, которые образовывали отдельные части черепа и лица. Между костями имеются швы, которые в молодом возрасте заполнены плотной соединительной тканью. Между некоторыми костями в онтогенезе образуются значительные соединительнотканные пространства, называемые родничками; к старости они окостеневают. Так, у млекопитающих такие роднички располагаются между затылочной и теменными костями черепа или между затылочной, теменными и височными.
Общая характеристика развития головного мозга
Уже давно подозревают, что дельфины имеют возможность распознавать людей и объекты, запоминать задачи, решать проблемы, адаптироваться к изменениям и изучать сложные задачи. Институт дельфинов в Гонолулу, Гавайи, проводит исследования поведения морских млекопитающих в течение последних 30 лет. Они значительно продвинули наше знание поведения дельфинов и интеллекта благодаря их работе. Ниже приведен краткий обзор результатов ряда исследовательских проектов.
Общая характеристика строения головного мозга
Дельфины, подобно людям, способны к поведенческой мимике. Другими словами, они могут имитировать поведение, продемонстрированное их тренерами-людьми. Если человек поднимает ногу, дельфин может распознать отношения, которые часть человеческого тела имеет к своей собственной анатомии, и поднимет свой хвост. Это указывает на то, что животное может ассоциировать часть своей анатомии с человеческой формой.
В процессе филогенеза некоторые кости, строящие скелет головы низших позвоночных, впоследствии меняют свои функции. Так, примитивные челюсти рыб заменяются покровными костями, но челюстные кости совсем не исчезают, они меняют свою функцию, оказываются у высших позвоночных слуховыми косточками среднего уха (рис. 34).
У наземных животных в связи со сменой жаберного дыхания на легочное на базе обонятельных ямок на лице развивается носовая полость, сообщающаяся в передней ее части с ротовой узким отверстием между резцовыми, верхнечелюстными костями и сошником. Эти отверстия были названы первичными хоанами. У животных, имеющих первичные хоаны, пища не может идти через ротовую полость тогда, когда через носовую полость проходит воздух. В связи с этим строение носовой полости совершенствуется и усложняется. Постепенно меняется расположение хоан, и уже у рептилий они открываются позади носовой полости в глотку и называются вторичными хоанами.
Он также может имитировать другого дельфина. Об этом свидетельствует производительность синхронного поведения. Если один дельфин собирается совершить лук, другое животное может скопировать это поведение и одновременно прыгнуть с этим животным. Говорят, что один дельфин действует как «демонстратор», в то время как другое животное является «подражателем» такого поведения.
Дельфины также могут интерпретировать телевизионные поведения и реагировать на жесты, показанные на экране, впервые подвергаясь воздействию телевидения. Это первая демонстрация любого вида животных поведенческого ответа на телевизионные жесты. Собаки, кошки и шимпанзе не показали таких ответов в аналогичных исследовательских протоколах. Это исследование изменило то, как мы в прошлом классифицировали дельфинов в первую очередь как акустические специалисты. Теперь мы понимаем, что они также являются визуальными специалистами, используя как зрение, так и звук, чтобы преуспеть в своей водной среде.
Череп связан с первым шейным позвонком (атлантом) и образует черепную полость, в которой помещается головной мозг. Эта черепная полость - cavum cranii имеет- крышу, заднюю, переднюю, боковые стенки и дно черепной полости.
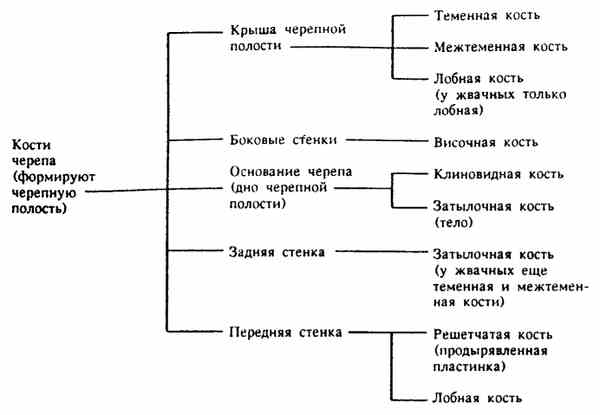
КРЫША ЧЕРЕПА - calvaria.
У свиней, лошадей и собак ее формируют парная лобная кость - os frontale (впереди), парная теменная кость - os parietale и непарная межтеменная кость - os interparietale- (сзади). Теменная кость граничит каудально с затылочной костью. У молодых животных на месте швов между теменной и затылочными костями образуется затылочный родничок, в нем закладывается парный очажок окостенения, из которого формируется непарная межтеменная кость. Она расположена по срединной линии в углу между двумя теменными и затылочной костями. Ее можно увидеть с внутренней стороны черепной полости в виде значительного костного утолщения, от которого направо и налево отходят костные гребешки. Это костный намет --tentorium osseum, к которому прикрепляется перепончатый намет твердой мозговой оболочки.
Было показано, что дельфины распознают себя в зеркале, используя так называемое «тестирование на случай непредвиденных обстоятельств» или делая движения, исследуя себя для этого движения. Это открытие неожиданно, поскольку дельфины в первую очередь испытывают мир через звук, и их система эхолокации не будет функционировать в двумерном отражении.
Концевой мозг- telencephalon
Они знают о своем собственном недавнем поведении и могут повторять поведение или, когда его спрашивает тренер, выполнять поведение, которое не было выполнено в последнее время. Команды, представляющие «повтор» или команды, представляющие «любые», приводят к повторению недавнего поведения или выбору какого-либо не-недавнего поведения соответственно. Это показывает способность дельфина поддерживать мысленный образ поведения, который он выполнял в последний раз, и обновлять это изображение по мере того, как выполняется каждое новое поведение, повторяя последнее поведение в этой последовательности по запросу.
У лошадей, свиней и собак переднюю часть крыши черепной полости образует лобная кость - os frontale. Она парная, представлена тремя пластинами:
1) верхней лобной, к которой впереди прикрепляется носовая кость;
2) орбитальной, опускающейся латерально и участвующей вместе с крыльями клиновидной кости и чешуей височной кости в формировании снаружи черепа внутренней стенки орбиты. (На стыке этих двух пластин снаружи от лобной кости отходит, образуя край орбитального кольца, скуловой отросток - processus zygomatics, который у лошадей и жвачных доходит до скулового отростка височной кости или височного отростка скуловой кости и образует полное кольцо орбиты, а у свиней и собак не доходит и образует неполное кольцо. У основания этого отростка можно встретить надглазничное отверстие - foramen supraorbitals.
Дельфины отвечают тренеру, указывающему на объект. Дельфины не только понимают и адекватно реагируют на человека, указывающего прямо на объект, они соответствующим образом реагируют на точку пересечения. Примером соответствующего ответа на указание было бы получить объект, на который указывает тренер, или перемещать объект из точки А в точку В.
Подобное исследование способности дельфинов оставаться сосредоточенными на задачах, требующих длительной концентрации, дало интересные результаты. Это демонстрирует способность дельфина долго оставаться внимательным и быстро разграничивать критические и некритические изображения и звуки с с высокой степенью точности.
У жвачных надглазничное отверстие расположено у основания скулового отростка, от него вперед и назад идет желоб, переходящий в сторону глазницы в канал.
У свиней отверстие переходит в желоб, достигающий носозых костей.
У собак надглазничного отверстия нет.);
3) внутренней вертикальной (не видной снаружи), образующей переднюю часть крыши черепной полости.
Помимо осознания себя, дельфины испытывают основные эмоции, участвуют в определенной степени абстрактной, концептуальной мысли, выбирают свои действия, учатся, наблюдая, понимают структуру своей среды, учатся тому, что работает, а что нет, путем решения проблем. создавать новые решения проблем, с которыми они представлены. При взаимодействии с человеком они, по-видимому, осознают разницу между детьми и взрослыми и склонны быть более нежными и терпеливыми с детьми.
Нейронаука предполагает, что человеческий мозг является стандартом, которому должны сравниваться мозги всех животных. Но многие животные демонстрируют уникальные таланты и очень продвинутые умственные способности и социальное поведение. Их интеллект сильно отличается от человека и в некотором роде превосходит его. Многие направления исследований показывают уникальные животные мозги с уникальными талантами в обновлении животных.
Между этими тремя пластинами (лобной, орбитальной и внутренней) в правой и левой лобных костях образуются пространства - полости, заполненные воздухом и выстланные слизистой оболочкой. Их называют пазухами или синусами. В лобной кости это парный лобный синус - sinus frontalis. Симметричные синусы между собой" разделены сагиттальной перегородкой и не сообщаются. Внутри пазух имеются еще несплошные перегородки, разделяющие каждую пазуху на ячейки, сообщающиеся между собой. Лобные пазухи переходят в теменные и затылочные (у свиней и крупного рогатого скота).
Известная философская статья спросила, можем ли мы понять, каково это быть летучей мышью. На самом деле, людям невозможно поместить себя в опыт существ с совершенно разными сенсорными и двигательными системами. Структуры в мозге животных совершенно разные, чем люди, и оценка их функции по сравнению с мозгом человека не объясняет очень продвинутого поведения.
Большинство животных обладают гораздо большим интеллектом, чем можно было бы ожидать путем простого анатомического сравнения с мозгом человека. В настоящее время хорошо известны интеллект слонов, дельфинов, китов, осьминог и приматов. Недавно было обнаружено, что дельфины вспоминают вокализации своих товарищей по танкам с двадцати лет до этого - невероятный подвиг памяти. Продвинутым интеллектом терапевтических собак было открытие, что многие животные имеют культуру.
У жвачных значительная особенность черепа в том, что крышу черепной полости формирует только лобная кость.
У крупного рогатого скота она имеет значительные размеры, внутри довольно мощные синусы, переходящие в теменные и носовые пазухи, и от наружной ее поверхности отходят еще полые роговые отростки - processus cornuales, полость которых сообщается с лобными синусами. Поэтому надо помнить, что при спиливании рога открывается вход в лобный синус через образовавшееся отверстие полого рогового отростка лобной кости. Задняя часть свода черепа формирует утолщенный край (межроговое возвышение) к которому снизу присоединяются теменные и межтеменная кости. Он подобен затылочному гребню у других животных. Лобная кость, образуя всю крышу черепной полости у крупного рогатого скота, как бы отодвигает на заднюю и боковую стенку полости теменные и межтеменную кости, которые у этих животных расположены сзади, несколько выше затылочной кости, под лобным гребнем. Теменные кости заходят на боковую стенку между лобной (сверху) и височной (снизу) костями, приобретая форму буквы П, сверху которой лобная кость, снизу (сзади) затылочная, а по бокам (снизу) височные кости.
Венозные синусы головного мозга
Этот пост будет посвящен нескольким маленьким мозгам мозга с различными структурами мозга, чем люди, но исключительные разведчики-птицы, рептилии и пчелы. Есть много других возможных примеров очень маленьких животных с интеллектом, таких как муравьи и термиты, но среди насекомых пчелы изучены больше всего. Сообщение завершится намеками относительно того, как такие маленькие мозги могут иметь расширенные возможности с структурами мозга, которые отличаются от людей.
Птицам нужен относительно большой мозг для их размера, чтобы летать. Но, по сравнению с людьми и другими крупными животными, их мозг очень мал. Совсем недавно было отмечено, что у какаду необычайная способность использовать многоступенчатые методы для разблокировки очень сложной клетки. Чтобы извлечь гайку, птица сначала удалила штифт, затем сняла винт, затем сняла болт, затем повернула колесо на 90 градусов, а затем сдвинула защелку сбоку. Эти птицы были неподготовленными и смогли выяснить, как это сделать менее чем за два часа.
У овец в отличие от крупного рогатого скота часть теменных костей участвует в образовании задней части крыши черепной полости и, направляясь на затылочную область, образует гребень теменной кости.
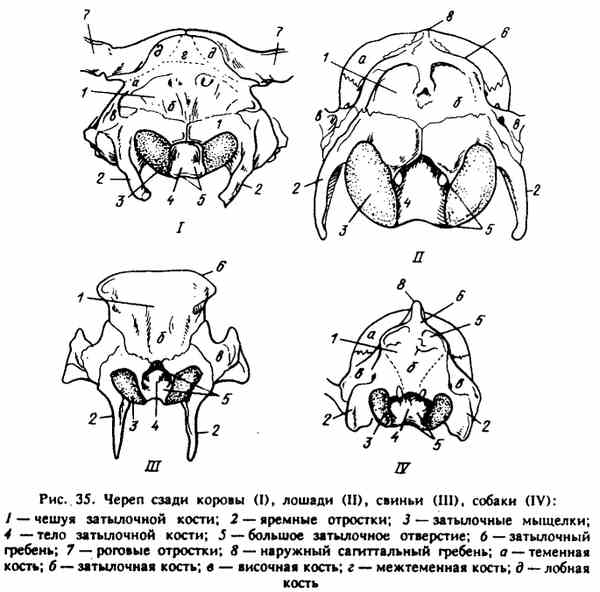
Рис. 35. Череп сзади коровы (I), лошади (II), свиньи (III), собаки (IV)
Артерии головного мозга
Другие могли учиться, наблюдая. Кроме того, как только они смогли открыть один из замков, они никогда не забыли, как и могли сделать это немедленно. Кроме того, какаду недавно полностью опровергли представление о том, что животные не могут танцевать, и, кроме того, они отмечают постоянство объекта на уровне четырехлетнего человека или обезьяны.
Джейс, наряду с попугаями, певчих птиц и ворон, являются одними из самых умных птиц. В последнее время сообщалось, что целая стая сойков наблюдалась в трауре погибшего товарища в течение 48 часов без какого-либо кормления. Каледонские ворон продемонстрировали выдающуюся память, в том числе воспоминание людей и автомобилей, которые раздражали их спустя годы. Они демонстрируют метапознание и подсчет, а также делают и используют передовые инструменты лучше, чем многие приматы. Они модернизировали инструменты, формовая кусок проволоки в крючок, чтобы получить труднодоступное удовольствие.
Характерно для костей черепа то, что с внутренней стороны черепной полости кости имеют гладкую поверхность, на которой видны небольшие углубления - вмятины, так называемые пальцевые вдавления - impressiones digitatae. Это отпечатки мозга
ЗАДНЯЯ СТЕНКА ЧЕРЕПНОЙ ПОЛОСТИ (рис. 35). Ее формирует затылочная кость - os occipitale. Oнa имеет три очага энхонд-рального окостенения, поэтому у молодняка подразделяется на три части: чешую - squamaoccipitalis, мыщелковые (боковые) части - partes laterales и тело затылочной кости (основная часть) - pars basilaris. Боковые (мыщелковые) части окружают большое отверстие - foramen magnum для выхода спинного мозга в позвоночный канал. При сильном наклоне головы вниз между затылочной костью и атлантом образуется пространство - доступ к спинному мозгу, что используется при убое животных. По бокам большого отверстия расположены округлые, покрытые суставным хрящом мыщелки затылочной кости - condyli occipitales, которые вместе с суставными ямками атланта образуют атлантозатылочный сустав.
Латеральнее мыщелков расположены яремные отростки - processus jugulares. Они очень мощные, прямые у свиней, несколько меньше у лошадей и жвачных и небольшие, чуть-чуть округлые у собак.
У основания яремных отростков медиально лежит круглое подъязычное отверстие - foramen hypoglossi, идущее в подъязычный канал, через который выходит из черепной коробки последняя - двенадцатая пара черепных нервов - подъязычный нерв - п. hypo-glossus.
У взрослого животного все три части затылочной кости срастаются. Чешуя затылочной кости у лошадей, свиней и собак расположена над мыщелковой частью, и верхняя ее часть образует мощный затылочный (выйный) гребень - crista occipitalis nuche). Он треугольный у с о б а к и образует прямоугольник у свиней.
ДНО ЧЕРЕПНОЙ ПОЛОСТИ (рис. 36). Его образуют основная часть (тело) затылочной кости и клиновидная кость. По краю тела затылочной кости образуется рваное отверстие - foramen lacerum (иногда в виде щели - у свиней и лошадей), через которое выходят черепные нервы (ветви V, IX, X и XI).
Клиновидная кость - os sphenoidale расположена впереди основания черепа. Если ее вычленить из черепа (в молодом возрасте, пока кости не срослись по швам), она имеет вид бабочки. Средняя/ непарная часть ее - тело - corpus sphenoidale расположено впереди от тела затылочной кости. По бокам от тела отходят две пары тонких костных пластин. Передние несколько больше задних - то крылья клиновидной кости - alae sphenoidales, вместе с теломгони формируют переднюю и среднюю черепные ямки, в которых лежат промежуточный и концевой мозг.
Обратите внимание на внутреннюю поверхность тела клиновидной кости, она имеет как бы две ступеньки: впереди верхняя - пресфеноид, сзади и ниже - базисфеноид, отделенные друг от друга узкой поперечной щелью, называемой желобом зрительного перекреста - sulcus chiasmatis. У молодых животных между ними синход-роз. На нижней ступеньке - базисфеноиде видна ямка, переходящая сзади в возвышение (на границе с телом затылочной кости). Эти образования напоминают по форме азиатское седло, в связи с чем и получили название «турецкое седло» - sella turcica. В его ямке лежит гипофиз (железа внутренней секреции), впереди от которого в желобе зрительного перекрестка расположен перекрест зрительных нервов, несущих импульс от сетчатки глаза в мозг.
По переднему краю наружной поверхности крыльев клиновидной кости сверху вниз располагаются отверстия, через которые черепные нервы связывают головной мозг с органами головы, расположенными на переднем участке стенки орбиты. Самое верхнее - решетчатое отверстие - foramen ethmoidale (оно образовано еще лобной костью), ниже - зрительный канал - canalis opticum, под которым у лошади и собака располагается глазничная щель - fissura orbitalis и еще ниже круглое отверстие - foramen rotundum (у свиней и жвачных они объединены в круглоглазничное отверстие).
На наружной стороне клиновидной кости располагаются крыловидные отростки, обрамляющие хоаны. Они большие и пластинча-тыеукрупного рогатого скота и свиней. У лошадей и собак в основании этих отростков проходит крыловидный канал - canalis pterygoideus, в котором идут верхнечелюстная артерия и нерв.
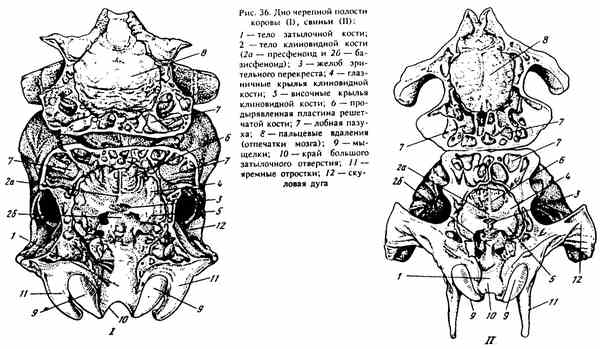
Рис. 36. Дно черепной полости коровы (I), свиньи (II)
БОКОВЫЕ СТЕНКИ ЧЕРЕПНОЙ ПОЛОСТИ. Сверху правые и левые парные крылья клиновидной кости швами соединяются с чешуей височной и лобной костей и формируют боковые стенки черепной полости. В молодом возрасте здесь имеется неокостеневший участок - клиновидный родничок - fonticulus sphenoidal.
Височная кость - os temporale участвует в формировании боковой стенки черепной полости. Она имеет четыре части: чешуйчатую часть - pars squamosa - пластину, формирующую боковую стенку; каменистую часть - pars petrosa - в каудальной части со стороны черепной полости; барабанную часть - pars tympanica - с наружной стороны черепа, внутри которой выделяется еще внутрибарабанная часть - pars endotympanica.
Латерально от наружной поверхности чешуи височной кости ответвляется скуловой отросток височной кости - processus zygomatics os temporale, который, изгибаясь, направляется вперед и соединяется с височным отростком скуловой кости, образуя вместе с ним скуловую дугу - arcus zygomaticus. На вентральной поверхности пластинчатого скулового отростка имеется расположенная в сегментальной плоскости суставная поверхность для соединения с нижней челюстью.
В передней своей части чешуя височной и крылья клиновидной костей образуют с внутренней стороны черепной полости боковые стенки, а снаружи - стенки орбиты, на поверхности которой и расположены по вертикали решетчатое отверстие, зрительный канал, глазничная щель и круглое отверстие, через которое выходят пять черепных нервов (II пара, III, IV, VI пары и ветви V пары), вследствие чего этот участок дна орбиты-один из самых насыщенных черепными нервами.
Каудальная часть чешуи височной кости прикрывает каменистую чйсть - pars petrosa (рис. 37), которая тоже участвует в образовании боковой стенки черепной полости и лежит впереди и выше от яремных отростков. Наиболее «самостоятельна» эта часть у л о ш а д и (она сравнительно легко отделяется), у других животных она очень прочно срастается с чешуей. Каменистая часть в основном предназначена для помещения в ней органов слуха и равновесия. Она видна со стороны черепной полости, плоская, гладкая, несет на себе отпечатки извилин мозга. Внутри нее помещается костный лабиринт - костные полости всевозможной формы, в которых расположены органы слуха и равновесия. Их этого лабиринта на поверхность каменистой части выходят две маленькие щели - наружные отверстия канальца улитки и водопровода преддверья - apertura externa vestibuli и apertura externa aqueductus cochlea.
Об этом обязательно надо помнить, так как именно через них полость костного лабиринта внутреннего уха сообщается с межоболочечными пространствами черепной полости; заболевание органов слуха в связи с этим может привести и к заболеванию мозговых оболочек - менингиту.
Со стороны черепной полости в средней части поверхности каменистой части расположено довольно крупное^ углубление, из которого в кость ведут два отверстия: внутренний слуховой проход - meatus acusticus internus, через который орган слуха и равновесия связан нервом (VIII) с продолговатым мозгом; отверстие для лицевого нерва (VII), который связывает мозг с лицевыми мышцами.
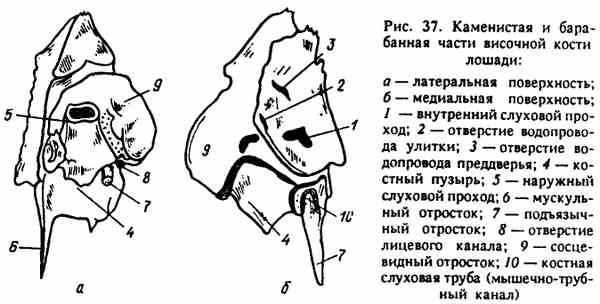
Рис. 37. Каменистая и барабанная части височной кости лошади
На внешней стороне каменистой части черепа снизу расположена барабанная часть - pars tympanica, а выше - сосцевидный отросток - processus mastoideus. На барабанной части хорошо заметен значительный (особенно у жвачных и свиней) барабанный пузырь - bulla tympanica. Внутри него полость - cavum tympani (место расположения среднего уха), в которую ведет вышерасположенная костная трубочка весьма значительного диаметра. Эта трубочка - наружный слуховой проход - meatus acusticus externus, проводящий звук, направляющийся из наружного уха через среднее во внут-реннее. Внизу у основания наружного слухового прохода хорошо виден (особенно у лошади) подъязычный (шиловидный) отросток - processus hyoideus (stylordeus), характерный тем, что имеет вид отточенной цилиндрической костной палочки. К подъязычному отростку прикрепляется подъязычная кость -os hyoideum, на которой подвешены язык, глотка и гортань. Здесь же виден острый мышечный отросток, оттянутый вниз, к которому прикрепляются мышцы нёбной занавески. У его основания видна большая щель, также ведущая в полость барабанного пузыря, где расположено среднее ухо. Эта щель - мышечно-трубный канал (слуховая труба) - canalis muscu-lotubarius (tuba auditiva), через который среднее ухо сообщается с полостью глотки. Это путь проникновения инфекции из глотки в среднее ухо.
Сосцевидный отросток - pocessus mastoideus довольно обширный, построен из губчатого вещества, покрытого тонким слоем компакты. Именно к нему прикладывает слуховой аппарат человек в том случае, если исключена возможность проведения звуковой волны через среднее ухо.
Под сосцевидным отростком расположено шиловидно-сосцевид-ное отверстие - foramen styJomastoideum, через которое дицевой нерв (VII) выходит из каменистой части для иннервации лицевых мышц.
Основание каменистой части на границе с телом затылочной кости может участвовать в образовании рваного отверстия.
ПЕРЕДНЯЯ СТЕНКА ЧЕРЕПНОЙ ПОЛОСТИ. Впереди от клиновидной кости, образуя переднюю стенку черепной полости, расположена решетчатая кость - os ethmoidale. Этa кость не видна на поверхности черепа. Она лежит на границе между черепной и носовой полостями. Ее можно видеть с внутренней стороны передней стенки черепной полости в виде двух округлых углублений, на дне которых видны пластинки с массой отверстий, это продырявленная пластина - lamina cribrosa. В отверстия этой пластины проходят к луковицам обонятельного мозга нервы, связывающие мозг с органом обоняния носовой полости.
Со стороны носовой полости к продырявленной пластине прикрепляется масса маленьких, различной величины, тончайших костных пластинок, образующих завитки. Эти наружные и внутренние турбиналии - эндотурбиналии и эктотурбиналии. В сумме эти завитки формируют лабиринт - labyrinths ethmoidalis. Правая и левая части лабиринта разделены довольно тонкой пластиной, расположенной по срединной сагиттальной линии, которая называется перпендикулярной пластиной. Со стороны черепной полости ее начало видно в виде вертикально поставленного гребешка, расположенного между двумя углублениями продырявленной пластины.
Со стороны носовой полости костная перпендикулярная пластина решетчатой кости продолжается вперед в виде уже хрящевой, никогда не окостеневающей носовой перегородки. От перпендикулярной пластины со стороны носовой полости направо и налево идут тонкие костные волнистые пластиночки, окружающие по бокам лабиринт. Получается, что основная часть, решетчатой кости - лабиринт, где расположен орган обоняния, лежит между тремя пластинами, образующими своеобразную «коробочку» с двумя отделениями (перпендикулярной пластиной эта коробочка разделена на два симметричных отделения). Задняя стенка коробочки (продырявленная пластина) видна со стороны черепной полости. Со стороны носовой полости на ней закреплен лабиринт. Эта коробочка открыта только со стороны носовой полости.
Чтобы хорошо усвоить расположение и название всех отверстий, через которые головной мозг, лежащий в черепной полости, сообщается с органами, к которым идут его нервы, посмотрите еще раз эти отверстия на черепе. На передней стенке через отверстия продырявленной пластины решетчатой кости черепная полость сообщается с лабиринтом решетчатой кости, т. е. с носовой полостью. На задней стенке через большое (затылочное) отверстие головной мозг переходит в спинной мозг, и черепная полость сообщается с позвоночным каналом. По бокам основания черепа, образованного телами затылочной и клиновидной костей, располагается серия отверстий, через которые черепные нервы связывают головной мозг с различными органами головы и тела. На наружной поверхности передней части стенки орбиты сверху вниз идет цепочка расположенных по вертикали отверстий: решетчатого, зрительного канала, глазничной щели и круглого, несколько каудальнее вдоль тела затылочной кости рваное отверстие, позади которого со стороны медиальной поверхности основания яремного отростка расположено круглой формы подъязычное отверстие (канал подъязычного нерва).
Оболочки головного мозга
Венозные синусы головного мозга
Артерии головного мозга
Общая характеристика строения головного мозга
Общая характеристика развития головного мозга
Концевой мозг- telencephalon
Развитие концевого мозга
Промежуточный мозг-diencephalon
Развитие промежуточного мозга
Средний мозг-mesencephalon
Развитие среднего мозга
Ромбовидный мозг-rhombencephalon
Развитие ромбовидного мозга
Список используемой литературы:
Оболочки головного мозга
Твердая оболочка головного мозга-dura mater encephali - самая наружная. В отличие от твердой оболочки спинного мозга она срастается с надкостницей черепных костей, поэтому эпидуральное пространство отсутствует. Между надкостницей и твердой мозговой оболочкой проходят лишь вены, образующие две системы венозных синусов - дорсальную и вентральную.
В субдуральное пространство от твердой мозговой оболочки отходят две складки: серповидная и перепончатый мозжечковый намет. Серповидная складка-falx cerebri- проходит сагиттально от петушьего гребня решетчатой кости до мозжечкового намета. Она лежит в продольной щели между полушариями большого мозга. Наиболее сильно серповидная складка выражена у собаки и лошади- она почти достигает мозолистого тела; она меньше у свиньи и коровы и особенно у мелких жвачных. Перепончатый мозжечковый намет-tentorium cerebella membranaceum- отходит оn костного намета или теменной кости, а также от гребней каменистых костей и располагается в поперечной щели между большим мозгом и мозжечком.
Паутинная оболочка головного мозга-arachnoidea encephali - на извилинах мозга прочно срастается с мягкой оболочкой. Подпаутинное пространство сохраняется только в щелях и бороздах между извилинами и на базальной поверхности мозга, где она образует расширения - вентральные цистерны продолговатого мозга, цистерну моста, червячка (и др.). Под паутинной оболочкой вдоль серповидной складки отходят в субдуральное пространство ворсинки, принимающие иногда форму узелков - пахионовы гранулы –granulationes arachnoidales. Пахионовы гранулы внедряются в сагиттальный синус и усиливают отток спинномозговой жидкости в вены.
Мягкая, или сосудистая, оболочка головного мозга-pia mater encephali - очень прочно срастается с мозгом, она заходит во все щели и углубления и вместе с сосудами проникает в вещество мозга. Вворачиваясь в полости мозга, она участвует в формировании сосудистых покрышек-tela chorioidea- в которых заложены сосудистые сплетения-plexus chorioideus. Такие образования находятся в желудочках мозга: боковых, третьем и четвертом.
Спинномозговая жидкость-liquor cerebrospinalis - заполняется субдуральное и субарахноидальное пространства головного и спинного мозга и через парные отверстия-apertura lateralis ventricularis quarti- каудально от боковых ножек мозжечка- и не парное отверстие-apertura mediana ventriculi quarti- сообщается с полостями желудочков и каналов мозга. Спинномозговая жидкость выделяется клетками эпендимы и сосудистых сплетений мозга. Она течет в субарахноидальном пространстве в сторону головного мозга, то есть краниально, а в центральном канале спинного мозга- каудально. Отток цереброспинальной жидкости происходит из субдурального пространства в венозную систему, а из субарахноидального пространства в органы лимфообращения.
Венозные синусы головного мозга
Дорсальная система синусов состоит из непарных сагиттального и прямого синуса и парных поперченных, затылочных и дорсальных каменистых; в них впадают вены мозга.
Сагиттальный синус-sinus sagittalis- находится в серповидной складке; назально он начинается из оболочечных вен, у лошади также из решетчатых и мозговых вен. В него впадают:
1)дорсальные вены мозга-vv.cerebri dorsales;
2)оболочечные вены из твердой мозговой оболочки;
3) костные вены-vv. diploicae;
4) короткий прямой синус-sinus rectus.
Последний образуется каудально от валика мозолистого тела, слиянием вены мозолистого тела с большой веной мозга.
Вена мозолистого тела-v. corporis callosi - выносит кровь из передней половины мозга.
Большая вены мозга-v. cerebri magna-принимает в себя глубокие вены мозга-vv. cerebri profundi- выносящие кровь из мозга и его сосудистых сплетений.
Каудально от впадения прямого синуса, сагиттальный синус делится на правый и левый поперечные синусы,-sinus transversus - каждый из которых направляется в височный канал и переходит в дорсальную мозговую вену-v. cerebri dorsalis. Последняя впадает в височную поверхностную вену. В поперечный синус, до его погружения в височный канал, вливается каменистый синус-sinus petrosus- который выносит кровь из базальных частей мозга.
Между поперечными синусами находится соединительный синус-sinus communicans- он лежит в основании мозжечкового намета. В него впадают затылочные синусы-sinus occipitales- помещающиеся в бороздах по обе стороны червячка мозжечка.
Отток крови из дорсальной системы синусов, помимо мозговых дорсальных вен, происходит так же и через эмиссарии-emissarium- в височные глубокие вены. Эмиссарии- это непостоянные отводящие каналы, прободающие стенку черепной полости.
Вентральная, или базилярная, система синусов включает синусы циркулярные и базилярные.
Циркулярный синус-sinus circularis-окружает гипофиз. Он образован правым и левым кавернозными синусами, соединяющимися межкавернозными синусами-sinus cavernosus et intercavernosus.
Каждый кавернозный синус назально переходит в глазничную вену мозга каудально- в базилярный синус. Глазничная вена мозга-v. cerebralis orbitalis- впадает через глазничную щель в глубокую лицевую вену.
Базилярный синус-sinus basilaris- вливается вентральный позвоночный синус. В области рваного отверстия он отдает вентральную и мозговую вену-v. cerebralis ventralis- которая впадает в затылочную вену. Близ затылочного сустава базилярные синусы, анастомозируя между собой, образуют вентральный затылочный синус-sinus occipitalis ventralis. В базилярную систему синусов открываются вентральные вены мозга.
Артерии головного мозга
1.Внутренняя сонная артерия-a. carotis interna -проникает в черепную полость через сонное или рваное отверстие и делится на назальную и каудальную соединительные ветви. С одноименными ветвями другой стороны они формируют вокруг гипофиза артериальное кольцо-circulus arteriosis cerebri. Назально из кольца выходит назальная артерия-a. cerebri nasalis- она направляется к мозолистому телу и питает передние отделы полушарий.
От назальной соединительной ветви ответвляются 4 сосуда:
а) назальная артерия мозговой оболочки-a. meningea nasalis;
б) средняя мозговая артерия-a. cerebri media-идет на боковую поверхность мозга;
в) назальная артерия сосудистого сплетения-a. chorioidea nasalis- по зрительному тракту следует в сосудистое сплетение бокового желудочка;
г) внутренняя глазничная артерия-a. ophthalmica interna- по зрительному нерву выходит в глазницу.
От каудальной соединительной ветви отходят:
а) каудальная мозговая артерия-a. cerebri caudalis- для задних отделов полушарий и четверохолмия
б) каудальная артерия сосудистого сплетения-a. chorioidea caudalis-в сосудистое сплетение боковых желудочков.
2. От затылочной артерии отходит спинномозговая артерия-a. cerebrospinalis- через межпозвоночное отверстие атланта она проникает в позвоночные канал и делится на краниальную и каудальную ветви, соединяющиеся с одноименными ветвями другой стороны. От краниальных ветвей в головной мозг отделяется основная артерия мозга-a. basilaris cerebri- она впадает в артериальное кольцо, а на своем пути отдает: позади моста- каудальную артерия мозжечка, впереди моста- краниальную артерию мозжечка-a. cerebella caudalis et cranialis- а к слуховому нерву- внутреннюю слуховую артерию-a. auditiva interna. Каудальная ветвь спинномозговой артерии анастомозирует со спинномозговой вентральной артерией-a. spinalis ventralis. Сосуды мозга иннервируются симпатической нервной системой.
Общая характеристика строения головного мозга
Головной мозг- encephalon- с дорсальной поверхности разделяется поперечной щелью-fissura transversa cerebri- на большой и ромбовидный мозг.
Большой мозг-cerebrum-состоит из двух полушарий, промежуточного и среднего мозга. Правое и левое полушария большого мозга-hemisphaerum cerebri dextrum et sinistrum- дорсально разграничиваются глубокой продольной щелью-fissura longitudinalis cerebri. Промежуточный и средний мозг дорсально прикрыты полушариями.
Ромбовидный мозг-rhombencephalon-состоит из мозжечка, мозгового моста, объединяемых в задний мозг, и продолговатого мозга.
Мозжечок-cerebellum- располагается дорсально от продолговатого мозга и сзади от полушарий большого мозга.
Продолговатый мозг-medulla oblongata- является непосредственным продолжением спинного мозга. На переднем конце его расположен мозговой мост-pons cerebri.
Впереди моста находятся ножки большого моста. Спереди ножки простираются до зрительных трактов-tractus opticus- и перекреста зрительных нервов-chiasma opticum. Между ножками большого мозга, позади зрительного перекреста, лежат: сосцевидное тело-corpus mamillare- и серый бугор с воронкой и гипофизом. Впереди зрительных трактов и с боков от ножек большого мозга выступает обонятельный мозг-rhinencephalon- орально он заканчивается двумя обонятельными луковицами-bulbi olfactorii. Обонятельный мозг представляет вентральный отдел полушарий большого мозга, а покрытый бороздами и извилинами плащ-pallium- дорсо-латеральный их отдел.
Полушария относятся к концевому мозгу-telencephalon- а зрительный тракты, серый бугор, воронка, гипофиз и сосцевидное тело - к вентральным отделам промежуточного мозга-diencephalon.
Ножки большого мозга являются вентральной частью среднего мозга-mesenecephalon.
Мозжечок и мост входят в состав заднего мозга-metencephalon. Промежуточный, продолговатый и средний мозг объединяются в ствол большого мозга-truncus cerebri.
Общая характеристика развития головного мозга
Головной мозг развивается в связи с возникновением и дальнейшим развитием различных высших нервных центров: рецепторных, моторных и ассоциативных, которые обеспечивают согласованную функцию всех органов - соматических, висцеральных, сосудистых.
На ранних стадиях онтогенеза и филогенеза головной мозг представляет собой расширенный конец мозговой трубки, который лежит впереди хорды, поэтому он называется прехордальным или первичным мозговым пузырем. Развитие его связано с органом обоняния, и его обонятельная функция сохраняется у всех животных, включая млекопитающих.
Несколько позже на переднем конце эпихордального мозга обособляется прехордальный, или вторичный, мозговой пузырь. Развитие его обусловлено: возникновением первичного ассоциативного и комиссурального центра; наличием жаберного аппарата и органов боковой линии водных животных; дифференцировкой внутренних органов.
От передней части эпихордального мозгового пузыря обособляется средний мозговой пузырь, который, исключая млекопитающих, выполняет ведущую роль, поскольку в него поступают импульсы решительно от всех рецепторных аппаратов и прежде всего зрительных.
С дорсальной поверхности все три отдела мозга довольно резко отделяются друг от друга поперечными комиссурами из нервных волокон, впереди и позади среднего мозга.
В дальнейшем и прехордальный мозг разделяется на концевой мозг в виде парного пузыря и промежуточный мозг.
Концевой мозг наивысшей стадии своего развития достигает у млекопитающих животных с наличием новой коры большого мозга, которая становится высшим нервным центром.
В промежуточном мозге у низших животных в связи с ведущей ролью обонятельного анализатора первое место занимают гипоталамус и эпиталамус. Они являются промежуточными обонятельными центрами, посылающими импульсы в средний мозг, а гипоталамус, кроме того, посылает импульсы в гипофиз и в автономные центры среднего и продолговатого мозга. В силу этого гипоталамус становится высшим подкорковым вегетативным центром. Эту функцию он охраняет и у млекопитающих животных.
Лишь с формированием коры большого мозга у млекопитающих усиленно начинает развиваться средний участок промежуточного мозга - таламус, через который кора получает решительно все рецепторные импульсы с периферии.
Таким образом, исторически гипоталамус более связан с эпихордальным мозгом, а таламус с прехордальным.
Почти одновременно с делением прехордального мозга дифференцируется и эпихордальный мозг позади среднего мозгового пузыря на задний мозг и продолговатый мозг. Задний мозг до млекопитающих представлен одним лишь мозжечком, который является подкорковым центром корреляции мышечных движений для сохранения равновесия. Лишь у млекопитающих к мозжечку вследствие развития новой коры большого мозга добавляется мозговой мост, а сам мозжечок усиливается двумя полушариями мозжечка.
Полости первичных мозговых пузырей становятся в развитом мозге мозговыми желудочками. Из полости прехордального мозга в концевом мозге возникают парные боковые желудочки, а в промежуточном мозге- третий мозговой желудочек. Все три желудочка соединяются межжелудочковым отверстием. Желудочек среднего мозгового пузыря вследствие разрастания стенок последнего превращения в мозговой водопровод, а полость ромбовидного мозга становится четвертым мозговым желудочком, который сообщается с центральным спинномозговым каналом.
Концевой мозг- telencephalon
Состоит из двух полушарий большого мозга-hemisphaerum dextrum et sinistrum-, разделенных с дорсальной поверхности глубокой продольной щелью-fissura longitudinalis cerebri.
В каждом полушарии рассматривают плащ, обонятельный мозг, полосатые тела и боковые желудочки мозга. Плащ расположен в полушарии дорсо-лательно от желудочка, обонятельный мозг – вентро-медиально. Границей между плащом и обонятельным мозгом на вентральной поверхности мозга служит базальная пограничная борозда. Полосатое тело лежит в вентральной стенке полушария дорсально от частей обонятельного мозга.
А) Плащ -pallium
Состоит из серого и белого мозгового вещества. Серое мозговое вещество-substantia grisea- образует кору большого мозга-cortex cerebri- на его поверхности. На ней различают извилины мозга-girus - разделенные бороздами и щелями. На латеральной поверхности плаща видны три дугообразные извилины. Они окружают латеральную сильвиеву борозду-sulcus lateralis. Дугообразные извилины четко выражены у собаки, а у других животных каждая дуговая извилина подразделяется бороздами на части. На медиальной поверхности плаща в основном заметны две дугообразные извилины вокруг мозолистого тела, причем ближайшая к нему поясная извилина-girus cinguli- функционально связана с обонятельным мозгом.
Белое мозговое вещество плаща-substantia alba- располагается под корой плаща. Оно состоит из проводящих путей - ассоциативных, комиссуральных и проекционных.
а) ассоциативные волокна соединяют отдельные участки коры в пределах каждого полушария. Они разделяются на короткие волокна и длинные.
б) комиссуральные волокна соединяют участки, принадлежащие разным полушариям. Они формируют мозолистое тело-corpus callosum- самую крупную комиссуру головного мозга. Оно помещается между полушариями в глубине продольной щели. Различают ствол мозолистого тела-truncus corporis callosi-и два конца - передний и задний. Передний конец называется коленом мозолистого тела-genu corporis callosi- он загибается вентрально. Задний конец, или валик, мозолистого тела-splenium corporis callosi- срастается со сводом.
Комиссуральные волокна, выходящие из ствола мозолистого тела, образуют лучистость спайки-radiatio corporis callosi- она формирует дорсальную стенку бокового желудочка мозга и расходится в передний, боковой и задний отделы коры плаща.
в) проекционные волокна - соединяют кору плаща, как с отдельными частями ствола головного мозга, так и со спинным мозгом. Они образуют в полосатом теле внутреннюю капсулу. Функционально проекционные пути разделяются на эфферентные и афферентные.
Эфферентные проводящие пути выносят импульсы из коры полушарий большого мозга в разные отделы ствола мозга: большого, ромбовидного и спинного.
Афферентные проводящие пути приносят импульсы в кору полушарий из спинного и ромбовидного мозга. Они образуют заднюю часть внутренней капсулы.
В противоположность коре больших полушарий все серое вещество остальных отделов всей центральной нервной системы объединяется понятием «подкорка». Импульсы со всех частей тела сначала идут в разные отделы подкорки, включая зрительный бугор, а из последнего поступают в кору полушарий. Кора постоянно разлагает, расчленяет возбуждения, возникающие в рецепторных приборах, и в то же время объединяет, синтезирует действие выделенных анализом возбуждений в разнообразные комплексы. Это и представляет собой проявление аналитической и синтетической деятельности коры головного мозга. Более того, высший отдел нервной системы - кора большого мозга является (у человека) материальным субстратом высшей нервной деятельности: мышления, сознания, памяти, рассудка, воли, то есть всей психической деятельности, которая представляет собой результат физиологической деятельности коры головного мозга.
Б) Обонятельный мозг-rhinencephalon
Располагается в вентро - медиальном отделе каждого полушария большого мозга. Отдельные части его видны на базальной и медиальной поверхностях полушарий, а также на дне боковых желудочков мозга. На базальной поверхности полушарий располагаются обонятельные луковицы, обонятельные тракты и извилины, обонятельные треугольники и грушевидные доли. На медиальных поверхностях полушарий видны околообонятельное поле, извилины гиппокампа, поясные извилины и поверхность разреза назальной спайки, а на дне боковых желудочков мозга- хвостатые ядра, гиппокамп и свод. Поясничная извилина, гиппокамп, свод и миндалевидное ядро составляют лимбическую систему-pars limbica rhinencephalon.
1.Обонятельная луковица-bulbus olfactorius - парное образование в виде довольно плоского, вытянутого и загнутого дорсально мозгового отростка, который выдается за передний край полушария мозга в обонятельную ямку решетчатой кости. Дорсомедиальный отдел луковицы построен из серого мозгового вещества, а латеро - вентральный – из белого мозгового вещества. В луковице находится желудочек обонятельной луковицы-ventriculus bulbi olfactorii- он является продолжением бокового желудочка мозга. В обонятельную луковицу входит обонятельный нерв-n. olfactorius. Он содержит многочисленные пучки нервных волокон –fila olfactoria- направляющихся от обонятельных клеток слизистой оболочки носа к нервным клеткам луковицы. Таким образом, обонятельные луковицы являются первичными обонятельными центрами.
2. От нервных клеток обонятельной луковицы начинаются обонятельные проводящие пути . Они образуют белое мозговое вещество самой луковицы и обонятельные тракты - общий, медиальный и латеральный-tractus olfactorius communis, medialis et lateralis. Латеральный обонятельный тракт переходит на грушевидную долю, покрывая на всем своем пути латеральную обонятельную извилину-girus olfactorius lateralis. Медиальный обонятельный тракт достигает медиальной поверхности плаща, формируя околообонятельное поле-area parolfactoria. Обонятельные тракты ограничивают обонятельный треугольник-trigonum olfactorium- из серого вещества. Они проводят импульсы от обонятельной луковицы к клеткам вторичных обонятельных центров в обонятельных извилинах, обонятельных треугольниках, околообонятельных полях, в грушевидных долях, а также в гипоталамус и в средний мозг.
3.Грушевидная доля-lobus periformis - расположена медиально от латерального обонятельного тракта и каудально от обонятельного треугольника; медиально граничит с ножками большого мозга. Каудо-медиальной границей грушевидной доли является медиальная пограничная щель, или щель гиппокампа-fissura hippocampi. В грушевидной доле заключена полость, представляющая собой задний отдел бокового желудочка мозга.
Грушевидная доля каудально переходит без четкой границы в извилину гиппокампа, расположенную на медиальной поверхности полушария, сзади и латерально от щели гиппокампа.
Извилина гиппокампа дорсально продолжается в поясную извилину-girus cinguli. Последняя проходит непосредственно дорсально от мозолистого тела,и огибая его спереди, соединяется с околообонятельным полем и представляет собой часть коры плаща. Грушевидная доля является вторичным обонятельным центром.
4.Гиппокамп, или аммонов рог-hippocampus - своим дорсальным отделом образует дно бокового желудочка мозга, позади хвостатого ядра, от которого отделяются сосудистым сплетением бокового желудочка. Гиппокамп представляет собой складку коры головного мозга в области щели гиппокампа и грушевидной доли. Он серповидно изгибается латер0-каудально и вентрально и теряется в стенке грушевидной доли. Аммоновы рога лежат дорсально на зрительных буграх, будучи отделены от них сосудистым сплетением третьего мозгового желудочка. Являясь высшими ассоциационными подкорковыми обонятельными и вкусовыми центрами, аммоновы рога связаны с различными участками коры больших полушарий и подкорковыми ядрами. Проводящие пути их образуют свод и его производные.
5. Свод -fornix - содержит в своем составе проводящие пути, соединяющие гиппокамп с сосцевидным телом промежуточного мозга. Отдельные участки этого пучка волокон образуют желобоватый листок, кайму аммонов рога, ножки, столбы, тело и переднюю спайку мозга.
Желобоватый листок-alveus hippocampi- покрывает гиппокамп с его поверхности, обращенной в боковой желудочек мозга. Он образован нервными волокнами, происходящими из серого мозгового вещества грушевидной доли и гиппокампа. По дорсолатеральному краю эти волокна создают кайму гиппокампа-fimbria hippocampi- продолжающуюся назально в ножку свода-crus fornicis. Последняя, соединяясь с ножкой с другой стороны, становится коротким телом свода-corpus fornicis. Тело свода служит дорсальной стенкой третьего мозгового желудочка; назально оно разделяется на два столба свода-columnae fornicis. Последние медиально от хвостатых ядер направляются к сосцевидному телу и серому бугру гипоталамуса. Комиссура свода-comissura fornicis- образована поперечными волокнами между ножками свода; она соединяет дорсальные концы аммоновых рогов друг с другом.
6. Между передним концом мозолистого тела и столбами свода находится передняя спайка мозга-comissura nasalis. Она соединяет друг с другом обонятельный мозг обоих полушарий.
В) Полосатое тело-corpus striatum
Расположено впереди зрительных бугров, дорсально от обонятельных треугольников, состоит из четырех ядер: хвостатого, чечевицеобразного, миндалевидного и ограды. Между ядрами находятся внутренняя и наружная спайка капсулы, образованные проекционными проводящими путями.
На хвостатом ядре-nucleus caudatus- различают головку и хвост. Головка хвостатого ядра своей дорсальной поверхностью образует дно бокового желудочка, назо -латерально от гиппокампа.
Чечевицеобразное ядро-nucleus lentiformis- располагается латерально от головки хвостатого ядра и зрительного бугра и отделяется о них внутренней капсулой-capsula interna. Латеральная часть чечевицеобразного ядра называется скорлупой-putamen. Медиальная часть чечевицеобразного ядра более древнего происхождения, называется бледным ядром-globus pallidus. Оно связано с обонятельным мозгом, лежит латерально от зрительного бугра и впереди от ядра латерального коленчатого тела-ganglion geniculatum. Латерально от чечевицеобразного ядра находится наружная капсула-capsula externa- а латерально от капсулы -другое ядро виде узкой полоски -ограда-claustrum. На границе между оградой, скорлупой и аммоновым рогом располагается миндалевидное ядро-nucleus amigdale- которое является обонятельным мозгом.
Полосатые тела соединяются проводящими путями: 1) с корой большого мозга; 2) со зрительными буграми и гипоталамусом; 3)с ядрами сетчатого образования среднего мозга, с ядрами моста и продолговатого мозга; 4) с ядрами черепно-мозговых нервов.
Через полосатые тела замыкаются различные рефлекторные цепи или:
А) периферический рецепторный аппарат - зрительные бугры - полосатые тела - соматические и висцеральные эффекторные аппараты.
Б) кора - полосатое тело - соматические и висцеральные эффекторные аппараты.
У млекопитающих ядра полосатых тел являются важнейшими подкорковыми двигательными центрами:
1) Координированных непроизвольных движений;
2) Регуляции мышечного тонуса в состоянии покоя и движения;
3) Безусловных рефлексов;
4) Высшими подкорковыми вегетативными центрами
Полосатые тела функционируют как единое целое, но отдельные их части действуют противоположно.
Развитие концевого мозга
Из более древних передней и вентральной стенок концевого мозга происходят обонятельные доли и полосатые тела, а из более поздней дорсальной его стенки - плащ.
1. Развитие обонятельных долей мозга обуславливается наличием обонятельного анализатора, который у водных животных имеет исключительное значение при ориентации во внешней среде. Отростки чувствительных обонятельных клеток заканчиваются в обонятельных луковицах, составляющих часть обонятельных долей концевого мозга, проксимальные отделы которых формируют древнейшую обонятельную кору-palaeocortex- из серого мозгового вещества. У млекопитающих она образует обонятельные извилины и обонятельные треугольники-trigonum olfactorium- являющиеся вторичными обонятельными центрами.
2. В вентральной стенке концевого мозга, дорсально от обонятельной коры очень рано появляется крупноклеточный базальный ганглий-ganglion basale. Это высший двигательный центр. Базальный ганглий сохраняется и у млекопитающих животных в виде бледного ядра. Позднее у наземных животных разрастаются дополнительные мелкоклеточные ядра, образующие в целом -neostriatum- который состоит из скорлупы, а у млекопитающих еще хвостатого ядра. Кроме того, обособляются новые ядра - ограда и миндалевидное ядро. С появлением вторичного плаща сложный базальный ганглий пронизывается внутренней и наружной капсулами из проводящих путей, идущих в кору плаща и обратно. В силу этого и возник новый термин -полосатое тело.
3. В эволюции плаща наблюдаются две формации различной функции и строения, не считая мембранообразного примитивного плаща-pallium membranaceum- характерного для водных животных и состоящего из эпендимы. У наземных животных сначала возникает кора более простой, трехслойной, клеточной структуры. Из нее формируются высшие обонятельные центры: грушевидные доли, извилины гиппокампа и их производные. Все эти части объединяются понятием первичный плащ-arhipallium.
Первичный плащ характеризуется расположением серого мозгового вещества под белым мозговым веществом. Архипаллиум, также как и палэопаллиум, лучше всего выражен у макросматических животных-macrosmates- обладающих острым обонянием, в отличие о микросматов-microsmates- у которых орган обоняния менее развит.
У млекопитающих на границе между обонятельной долей и первичным плащом появляется вторичный плащ-neopallium. Вторичный плащ характеризуется поверхностным расположением серого мозгового вещества в виде коры над белым мозговым веществом. Зачатки его появляются еще у рептилий. По мере своего разрастания новый плащ оттесняется от арихипаллуим в медиальную сторону и вентрально. При этом в области щели гиппокампа образуется складка коры в виде гиппокампа, а сам неопаллиум занимает всю дорсолатеральную часть полушария и, распространяясь каудально, покрывает промежуточный и даже средний мозг с их дорсальной и боковых поверхностей.
Разрастание вторичного плаща вызывается внедрением в него новых проекционных нервных волокон из промежуточного мозга, являющихся проводниками различных анализаторов- кожного, зрительного, слухового, мышечного и развитием новых ассоциативных нейронов и центров, включая сетчатое образование.
Кора вторичного плаща в ряду животных чрезвычайно усложняется в своем строении, резко дифференцируются ее функции и структура и увеличиваются размеры. У крупных млекопитающих животных плащ обычно несет извилины и борозды. Такие животные называются-gyrencephali. Извилины в ряду животных располагаются неодинаково. В одних случаях они в основном идут дугами вокруг поперечной сильвиевой борозды; у приматов извилины образуют две системы - лобную и теменную. Обе системы разделены сильвиевой бороздой. У третьей группы животных сильвиева борозда отсутствует, а борозды идут в передней части мозга, продольно, а в задней поперечно. Поэтому гомологизация извилин между животными разных отрядов крайне затруднительна, а в отдельных случаях может быть и невозможна.
У мелких животных извилины вообще отсутствуют. Наибольшее число извилин насчитывается у слонов и китов. В онтогенезе извилины появляются также не сразу, а в определенной последовательности.
Промежуточный мозг-diencephalon
Располагается позади полосатых тел концевого мозга и впереди среднего мозга, дорсально прикрыт сосудистой покрышкой третьего мозгового желудочка и гиппокампом. Промежуточный мозг состоит из трех отделов различного происхождения, строения и функции: эпиталамуса, таламуса и гипоталамуса. Эпиталамус образован сосудистой покрышкой третьего мозгового желудочка, эпифизом и парным узлом уздечки. Таламус состоит из зрительных бугров, между которыми находится кольцевидной формы третий желудочек мозга. Гипоталамус составляют зрительный выступ с концевой пластинкой, серый бугор с воронкой и гипофизом и сосцевидное тело. Все части гипоталамуса видны на базальной поверхности мозга, между ножками большого мозга, позади перекреста зрительных нервов.
Таламус. Зрительные бугры-thalami optici- самая массивная часть промежуточного мозга. Назо-латерально они срастаются с хвостатыми ядрами полосатого тела; зрительные бугры отделятся от последних пограничной полоской-stria terminalis-, от четверохолмия - поперечной бороздой, а друг от друга - ямкой зрительных бугров, прикрытой сосудистой покрышкой третьего желудочка. Бугры составляют многочисленные скопления ядер серого мозгового вещества. Наиболее крупные из них:
1.назальное ядро-nucleus nasalis thalami - лежит в толще назального бугорка-tuberculum nasalis thalami- в назо-медиальном отделе зрительного бугра. Оно наиболее древнее и является центром переключения афферентных путей на рефлекторные пути.
2.каудальное ядро-nucleus caudalis thalami - заключено в толщу каудо-латерального отдела латерального бугорка-tuberculum lateralis thalami- и состоит из промежуточных зрительных и слуховых центров. От перекреста зрительных нервов и слуховых центров. От перекреста зрительных нервов-chiasma opticum- на базальной поверхности мозга начинаются зрительные тракты-tractus opticum. Каждый тракт огибает латерально таламус и переходит в латеральное коленчатое тело-corpus geniculatum lateralis- теряющееся в каудальном ядре зрительных путей, идущих в кору большого мозга. Между латеральным коленчатым телом и четверохолмием выступает медиальное коленчатое тело-corpus geniculatum medialis. Оно соединяет каудальные холмы четверохолмия с каудальным ядром зрительного бугра и является промежуточным слуховым центром на пути в кору. Оба коленчатых тела объединяются в 7.
3. латеральное ядро-nucleus lateralis thalami - является центром переключения проводящих путей кожного анализатора и суставно-мышечного анализатора на проводящие пути, идущие в кору большого мозга и в полосатое тело.
4. медиальное ядро-nucleus medialis thalami - представляет собой промежуточный двигательный центр для проводящих путей из коры в ядра экстрапирамидной системы: в красное ядро, в ядра черепно-мозговых нервов и в спинной мозг.
5. сетчатое образование-formatio reticularis - располагается между ядрами и связано с ними.
Третий желудочек мозга -ventriculus tertius-находится между зрительными буграми, имеет кольцевидную форму, так как в него прорастает промежуточная масса зрительных бугров-massa intermedia thalami. В стенках желудочка находится центральное серое мозговое вещество-substantia grisea centralis- в нем располагаются подкорковые вегетативные центры. Третий желудочек сообщается с мозговым водопроводом среднего мозга, а позади назальной спайки мозга-comissura nasalis- с боковыми желудочками мозга через межжелудочковое отверстие-foramen interventriculare.
Эпиталамус . По краям ямки зрительных бугров видны мозговые полоски зрительных бугров-stria medullaris- а на них – парный узел уздечки-ganglion habenula. Узел уздечки переходит в уздечку или поводок-habenula- на нем укреплен эпифиз-epiphysis- грушевидной формы. Эпифиз-железа внутренней секреции, лежит в ямке между зрительными буграми и четверохолмием. Ганглий уздечки служит промежуточным центром для рефлекторных путей между мозгом, ядрами 5 пары и межножковым ядром.
Сосудистая покрышка третьего мозгового желудочка-tela chorioidea ventriculi tertii - образована складкой эпителиальной пластинки мягкой оболочки мозга и сосудистым сплетением. Эпителиальные пластинки покрышки прикрепляются по краям ямки зрительных бугров и свода. Сосудистая покрышка отделяет зрительные бугры от аммоновых рогов и от свода, она проникает через межжелудочковое отверстие в боковые желудочки мозга в виде сосудистых сплетений боковых желудочков мозга-plexus chorioideus ventriculi lateralis. Сосудистая покрышка формирует впереди эпифиза и непосредственно позади валика мозолистого тела выступ-recessus suprapinealis.
Гипоталамус - подталамическая, более древняя часть-hypothalamus- образует стенку третьего желудочка вентрально от зрительных бугров, является высшим подкорковым вегетативным центром, соединяется со зрительными буграми, с обонятельным мозгом и со средним мозгом. Гипоталамус содержит центры регуляции: дыхания, кроволимфообращения, температуры, обмена веществ, половых функций. Передний отдел его представлен серым бугром, воронкой и гипофизом, а задний- сосцевидным телом и стенками третьего желудочка.
Серый бугор гипоталамуса-tuber cinereum - лежит непосредственно позади перекреста зрительных нервов, между ножками большого мозга. В центра серого бугра имеется бухта воронки-recessus infundibuli. Сама воронка-infundibulum- тонкостенная, к ней прикрепляется гипофиз.
Гипофиз-hypophysis - представляет собой плоскоокруглое тело сложного строения с небольшой центральной полостью. Гипофиз состоит из трех частей: мозговой, промежуточной и железистой. Он является важнейшей железой внутренней секреции, выделяющей разнообразные гормоны и регулирующей все остальные железы внутренней секреции, симпатическую и парасимпатическую нервную системы.
Сосцевидное тело-corpus mamillare - лежит непосредственно позади серого бугра и служит промежуточным рефлекторным обонятельным центром, который через комплекс образований свода соединяется с обонятельным мозгом. Кроме того, сосцевидное тело связано со зрительными буграми и с сетчатым образованием среднего мозга. У собаки сосцевидное тело парное.
Развитие промежуточного мозга
Промежуточный мозг закладывается из небольшого числа клеток в стенке обширного третьего желудочка мозга, лишь у наземных животных. И особенно у млекопитающих, он достигает значительного размера.
1.хороо выраженная эмбриональная пластинка покрышки у всех взрослых млекопитающих формирует эпиталамус. Эпифиз является рудиментом третьего непарного теменного глаза. Лишь у некоторых водных животных и у рептилий имеется глазообразный пузырь под кожным покровом. У млекопитающих эпифиз становится железой внутренней секреции, функция которой еще недостаточно изучена.
2.своеобразно и сильно развитая эмбриональная пластинка дна формирует подталамическую часть промежуточного мозга. Она происходит из эпихордального мозга, то есть связана со средним мозгом.
Гипофиз происходит из трех разных источников. Эктодерма глотки превращается в разветвленную железу. Просвет железы впоследствии исчезает, но остаются тяжи из железистых клеток разнообразного строения, окруженные большим числом сосудов. У наземных животных из стенки воронки возникает нервная часть гипофиза из нервных и глиальных клеток и нервных волокон. Обособляется промежуточная часть гипофиза из эпителиальных клеток. Гипофиз в целом выделяет свыше десятка разнообразных гормонов, которые из железистой части поступают в кровь, а из нервной и промежуточной частей - в спинномозговую жидкость. Гипофиз находится во взаимодействии с вегетативными центрами, заложенными в стенках третьего желудочка мозга.
3.эмбрональная боковая пластинка образует зрительные буры-thalamus - и глазные пузыри. В состав таламуса входят ядра зрительных бугров. Они служат:
а) промежуточными центрами всех проводящих путей, которые направляются в кору полушарий и проводят различные импульсы - обонятельные, кожной и мышечной чувствительности, вкусовые, а у наземных животных, кроме того, зрительные и статоакустические;
б) промежуточным центром всех проводящих путей, идущих из коры плаща в разные отделы мозга.
Этим объясняется, почему зрительные бугры начинают формироваться с появлением обонятельного плаща и достигают максимального развития у млекопитающих в связи с образованием вторичного плаща. Мощное развитие зрительных бугров обусловлено также перемещением зрительных центров из среднего мозга во вторичный плащ, связью с мозжечком. В результате обогащения таламуса промежуточными центрами разрастается промежуточная масса зрительных бугров, которая внедряясь в полость третьего желудочка мозга, превращает его в кольцевидный канал. В сером веществе стенки третьего мозгового желудочка находятся многочисленные высшие подкорковые вегетативные центры.
Средний мозг-mesencephalon
Состоит из: пластинки четверохолмия, покрышки ножек, или чепца, из ножек большого мозга. Полость среднего мозга превратилась в мозговой водопровод-aqueductus cerebri- он соединяет третий и четвертый мозговые желудочки. В стенках водопровода заложено центральное серое мозговое вещество покрышки.
1.пластинка четверохолмия-tectum - представляет дорсальную часть среднего мозга, она лежит каудально от зрительных бугров и назально от мозжечка. Пластинка состоит из парных назальных и каудальных холмов-colliculi nasalis et caudalis. Холмы разделяются поперечной и срединной бороздами. С поверхности пластинка четверохолмия покрыта белым мозговым веществом, под которым находится серое мозговое вещество, в назальных холмах оно является подкорковым центром разнообразных афферентных путей, в то числе и зрительных, а в каудальных- подкорковым слуховым и равновесным центром.
В целом пластинка четверохолмия является координирующим центром ряда импульсов: обонятельных, зрительных, равновесно-слуховых, общей чувствительности и импульсов из коры больших полушарий. Двигательные импульсы передаются в красное ядро, в спинной мозг, а также к главным мышцам, в мозжечок и в мост.
2 .в центре среднего мозга между пластинкой четверохолмия и ножками большого мозга находится покрышка ножек или чепец-tegmentum pedunculi . В нем залегают парные ядра серого мозгового вещества, в плоскости передних холмов лежат:
а) красное ядро-nucleus ruder-двигательный центр спинного мозга;
б) ядро глазодвигательного нерва-nucleus n. oculomotorius.
в) парасимпатические ядра Якубовича.
Каудальнее располагаются:
г) ядро блокового нерва-nucleus n. trochlealis;
д) часть ядра пятой пары нервов.
Через весь чепец из продолговатого мозга в промежуточный проходит сетчатое образование, образующее проводящие пути из спинного мозга и мозжечка в четверохолмие, в зрительные бугры, и из них в спинной мозг.
3.ножки большого мозга-pedunculi cerebri - выступают на базальной поверхности мозга в виде двух толстых валиков между зрительными трактами и мозговым мостом. Они разделены межножковой бороздой. Из ножек выход 3 пара черепно-мозговых нервов - глазодвигательный нерв. Ножки образованы эфферентными проводящими путями, которые соединяют кору плаща с ромбовидным и спинным мозгом. Поэтому ножки сильнее развиты у тех животных, у которых кора полушарий более мощная. У человека они развиты наиболее сильно.
Развитие среднего мозга
Средний мозг у низших животных, как и у эмбрионов высших животных достигает очень значительных размеров. Серое мозговое вещество эмбриональных боковых пластинок среднего мозгового пузыря, разрастаясь, формирует свод среднего мозга, из которого у низших животных происходит двухолмие, или зрительные доли. Первоначально двухолмие было высшим координирующим центром, поскольку в нем оканчиваются не только зрительные нервы, но и афферентные проводящие пути из полосатого тела, ромбовидного и спинного мозга. Но уже у рептилий часть волокон, а у млекопитающих почти все волокна зрительного нерва перемещаются через зрительные бугры промежуточного мозга в кору вторичного плаща. Поэтому зрительные доли отстают в росте, а вторичный плащ и соответственно зрительные бугры разрастаются все сильнее и сильнее.
У наземных животных в своде среднего мозга закладывается равновесно-слуховые центры, первоначально в виде микроскопических образований, а позднее и макроскопических. Лишь у млекопитающих вместо двухолмия появляется четверохолмие. У животных с хорошим слухом преобладают задние - слуховые холмы.
Из эмбриональной основной пластинки в вентральной стенке среднего мозгового пузыря формируется чепец. В результате образования свода и чепца полость среднего мозга превращается в мозговой водопровод. Чепец состоит из ядер 3 и 4 пар черепно-мозговых нервов и специальных двигательных ядер чепца. К последним относятся красное ядро, связывающее мозжечок со спинным мозгом, и межножковое ядро, соединяющееся через ганглий уздечки с обонятельным мозгом.
У млекопитающих в связи с развитием вторичного плаща чепец вентрально порывается слоем белого мозгового вещества из проводящих путей, идущих из коры в ромбовидный и спинной мозг. Эти проводящие пути формируют ножки большого мозга, мощность которых соответствует степени развития коры полушарий большого мозга.
Ромбовидный мозг-rhombencephalon
Подразделяют на продолговатый и задний мозг. Задний мозг состоит из мозжечка и мозгового моста. Между мозжечком и продолговатым мостом находится четвертый мозговой желудочек.
А) Продолговатый мозг-medulla oblongata
Каудально продолжается без заметной границы в спинной мозг. На базальной поверхности его хорошо видна вентральная срединная борозда –fissura mediana ventralis. По обе стороны от нее проходят боковые борозды-sulcus paramedianus- каудально они вливаются в срединную вентральную борозду. Между этими тремя бороздами выступают два узких валика- пирамиды-pyramis- в них проходят пирамидные проводящие пучки от коры большого мозга в спинной мозг. Так как проводящие пучки продолжаются в боковые канатики и спинного мозга и при этом перекрещиваются справа налево и наоборот, то образуется перекрест пирамид-decussatio pyramidum. Латерально от пирамиды позади моста выходит 4 пара черепно-мозговых нервов - отводящий нерв. Близ перекреста пирамид и латерально от него отходит 12 пара - подъязычный нерв, латерально же от подъязычного нерва один за другим еще два нерва: 10 пара- блуждающий нерв, более назально- 9 пара- языкоглоточный нерв.
Серое мозговое вещество продолговатого мозга сгруппировано:
а) в отдельные чувствительные и двигательные ядра, из которых выходят 5, 6,7,9,10 и 12 пары черепно-мозговых нервов;
б) в промежуточные ядра пучков Голля и Бурдаха;
в) в ядра 8 пары и связанные с ними назальные и каудальные оливы-7.
Среди ядер залегает сетчатое образование-formatio reticularis- из переплетающихся нервных волокон и нервных клеток между ними, которое из продолговатого мозга переходит в чепец среднего мозга и в промежуточный мозг. Оно выполняет, прежде всего, ассоциационную и координационную функцию между различными ядрами ромбовидного и среднего мозга и является центром дыхания и сердечнососудистой системы.
Белое мозговое вещество продолговатого мозга состоит из большого количества пучков проводящих путей, идущих из спинного мозга в различные отделы головного мозга и обратно.
Б) Мозжечок-cerebellum
Имеет почти шаровидную форму, двумя бороздами он разделяется на среднюю часть- червячок и две боковые доли. Серое мозговое вещество образует кору мозжечка-cortex cerebelli- и отдельные ядра, залегающие в центрально расположено белом веществе мозжечка. Функция- координация движений.
Поверхность коры червячка-vermis- изрезана поперечными бороздами и щелями. Двумя основными щелями червячок разграничен на переднюю, среднюю и заднюю доли-lobus rostralis, caudalis et medialis. Каждая из них связана с ножками мозжечка - передними, средними и задними, состоящими из проводящих путей. Передний и задний концы червяка загнуты вентрально и навстречу друг другу, между ними остается небольшая щель- верхушка шатра-fastigium. Белое мозговое вещество червячка-7- на сагиттальном разрезе напоминает ветку туи, поэтому оно получило название- древо жизни-arbor vitae. В белом мозговом веществе червячка находится шатровое ядро-nucleus fastigii- которое является подкорковым центром равновесного анализатора.
С продолговатым мозгом червячок соединяется через каудальный мозговой парус-velum medullare caudale- а с четверохолмием через назальный мозговой парус-velum medullare nasale.
Полушария мозжечка-hemisphaerum cerebelli- как и червячок состоят из многочисленных долек, из которых одна - клочок-flocculus- в виде небольшого придатка червячка лежит сзади от боковой ножки. В белом мозговом веществе полушарий мозжечка находятся зубчатые ядра-nucleus dentatus- служащие передаточными центрами двигательных импульсов.
Мозжечок соединяется с продолговатым мозгом. С мозговым мостом - боковыми ножками и со средним мозгом - назальными ножками.
Каудальные ножки мозжечка-pedunculus cerebellaris medullum oblongatum- в виде двух валиков выступают на дорсальной поверхности продолговатого мозга. В них проходят проводящие пути:
а) из спинного мозга- пучки Флексига, Голля и Бурдаха с их ядрами;
б) из ядер слухового нерва;
в) из каудальных олив;
г) из ядер 5,10 и 12 пар черепно-мозговых нервов.
Назальные ножки мозжечка-brachium conjum ctivum cerebelli- идут под задними холмами четверохолмия в ножки большого мозга. Назальные ножки лежат на дорсальной поверхности продолговатого мозга. В них проходят проводящие пути:
а) из спинного мозга в червячок- пучок Говерса;
б) из зубчатых ядер полушарий мозжечка в красное ядро;
в) в ядра зрительных бугров.
Собственные проводящие пути мозжечка представлены волокнами между корой мозжечка и его ядрами и ассоциационными волокнами, соединяющими друг с другом отдельные извилины в сагиттальной плоскости.
В) Мозговой мост-pons cerebri
Лежит на переднем конце продолговатого мозга, на границе его со средним мозгом, в виде поперечного валика, который своими концами загибается дорсально к мозжечку, образуя боковые ножки мозжечка-brachium pontis. Мост и ножки состоят из проводящих путей, соединяющих ядра моста-nucleu pontis- с ядрами мозжечка. В ядрах моста заканчиваются проводящие пути из коры большого мозга и начинаются проводящие пути в полушария мозжечка. Через боковые отделы моста выходит 5 пара - тройничный нерв - самый массивный нерв из всех черепно-мозговых нервов. Он имеет два корня: вентрально - назальный двигательный и дорсо - каудальный чувствительный. На последнем находится большой полулунный ганглий. Каудально от моста, также в поперечном направлении, лежит трапецоидное тело-corpus trapezoideum- в виде узкого и низкого валика. Оно сформировано проводящими путями, идущими от ядер слухового нерва. Через боковые части трапецоидного тела выходят 8 пара- равновесно-слуховой нерв и 7 пара -лицевой нерв.
Г) Четвертый мозговой желудочек-ventriculus quartus
Помещается между мозжечком и продолговатым мозгом. Сводом ему служат червячок и мозговые паруса, а дном - продолговатый мозг.
Дно четвертого желудочка называется ромбовидной ямкой-fossa rhomboidea. Срединной и двумя боковыми бороздами на дне ямки выделяется парное срединное возвышение-eminentia medialis- на котором против латеральной ножки мозжечок выступает лицевой холмик-colliculi facialis. В области лицевого холмика залегают ядра отводящего и лицевого нервов. На каудальном конце срединного возвышения находится поле подъязычного нерва-area hypoglossi- с одноименным ядром.
Латерально от подъязычного нерва выступают ядра 9 и 10 пар нервов. Они формируют серое крыло-ala cinerea. Область заднего конца серых крыльев известна под названием писчего пера-calamus scriptorius.
Непосредственно позади боковых ножек мозжечка и медиально от них выступают в виде небольших возвышений вестибулярные поля-area vestibularis. Они содержат вестибулярные и улитковые ядра 8 пары нервов. Улитковые ядра лежат латеральнее.
Развитие ромбовидного мозга
Продолговатый мозг развивается путем разрастания боковых и основных пластинок эмбриональной мозговой трубки. Вследствие раздвигания боковых пластинок формирующимся мозжечком двигательные центры продолговатого мозга оказались оттесненными медиально, а чувствительные латерально.
Эти же обусловливается и медиальное расположение на продолговатом мозге двигательных нервов и латеральное - чувствительных, а также особенности строения дна четвертого желудочка мозга, в толще которого чувствительные ядра лежат латерально от двигательных.
Строение продолговатого мозга в целом у животных макроскопически довольно однообразно, что объясняется и однообразной его функцией - центры жаберного аппарата и его производных, кишечника, центры органов осязания, суставно- мышечного чувства, слуха и равновесия. Ввиду того, что через продолговатый мозг проходят проводящие пути, соединяющие головной мозг со спинным, его серое мозговое вещество разбивается на отдельные ядра в отличие от компактного серого мозгового вещества спинного мозга.
В продолговатом мозге заслуживает особого внимания моторное ядро чепца или сетчатое образование, которое впервые возникает у рыб и является древнейшим ассоциационным и двигательным центром. У наземных животных за счет моторного ядра чепца формируются назальным и каудальные оливы как ассоциационные центры. Назальные оливы появляются только у наземных животных, начиная с амфибий. Они служат промежуточным центром на пути от улиткового нерва к зрительным буграм. Пучок этих волокон образует трапецоидное тело. Каудальные оливы формируются еще позднее у птиц и млекопитающих. Они связаны с ядрами дорсальных канатиков, со зрительными буграми, с мозжечком и спинным мозгом. По своим связям каудальные оливы имеют самое ближайшее отношение к функции сохранения равновесия.
Мозжечок развивается в связи с выполняемой им функцией сохранения равновесия тела и поддержания мышечного тонуса. Поэтому он выражен наиболее сильно у животных, быстро плавающих, бегающих, прыгающих, летающих, и слабее у животных, передвигающихся медленно. В примитивном виде мозжечок представляет непарную пластинку и бело или серого мозгового вещества. Мозжечок закладывается в средней части эмбриональной пластинки покрышки ромбовидного мозга, а из передней и задней частей последней формируются передний и задний мозговые паруса. Пластинка мозжечка, разрастаясь спереди назад, изгибается дугообразно в дорсальном направлении. Благодаря появлению продольных борозд обособляется средняя часть пластинки- тело мозжечка- и парные боковые части- ушки мозжечка. Тело мозжечка и ушки относятся к palaeocerebellum. Тело мозжечка у наземных животных разделяется поперечными бороздами на переднюю, среднюю и заднюю доли, на которых возникают добавочные поперечные борозды. Передняя доля связана с мышцами головы, а средняя и задняя- с мышцами туловища и конечностей.
У млекопитающих преобладает средняя доля. Продольные борозды на ней отделяют среднюю непарную часть- червячок - от парных боковых долей, или полушарий мозжечка. Полушария мозжечка относятся к neocerebellum. В червячке находятся центры координированных, синхронных движений туловища и конечностей. Полушария мозжечка сильнее всего выражены у высших млекопитающих, обладающих в большей или меньшей степени способностью обособленных движений конечностей. Совершенствование этой способности, в свою очередь зависит от мощности коры большого мозга, как высшего центра нервной деятельности, и от возникновения связей мозжечка с корой большого мозга через боковые его ножки и мост.
Полушария мозжечка и мост имеются только у млекопитающих и достигают максимального своего развития у приматов. Однородной функцией мозжечка у животных объясняется довольно однообразное гистологическое строение его коры, в которой различают поверхностный молекулярный слой, глубокий зернистый и средний слой из крупных клеток Пуркине.
Ушки примитивного мозжечка у водных животных имеют отношение к органам равновесия, то есть органам боковой линии, и к мускулатуре хвоста. С редукцией этих органов у наземных животных уменьшаются и ушки. У млекопитающих от них сохраняются клочки, соединяющиеся с задней долей червячка.
Онтогенез червячка также определен его функцией, что особенно хорошо видно на мозге человека. У новорожденного ребенка он еще очень мал и составляет лишь 1/16-1/18 веса большого мозга. Но эти соотношения изменяются по мере роста ребенка и увеличения его подвижности, у взрослого отношения уже иные – 1/8-1/9 веса мозга.
Список используемой литературы
1) А.И. Акаевский «Анатомия Домашних Животных»
2) И.В. Хрусталева «Анатомия домашних животных»
3) Л.В. Антипова, В.С. Слободяник, С.М.Сулейманов «Анатомия и гистология сельскохозяйственных животных»
4) А.Ф. Римов «Анатомия домашних животных»
5) А.Ф. Кузнецова «Справочник по ветеринарной медицине»