Descrieți principalele tipuri de țesut muscular. Structura histologică a țesutului muscular.
Țesutul muscular se numește țesut diferit în structură și origine, dar similar în capacitatea de contracții pronunțate. Acestea asigură mișcarea în spațiul întregului organism ca întreg sau părțile sale (de exemplu, mușchii scheletici) și mișcarea organelor în interiorul organului (de exemplu, inima, limba, intestinele).
Celulele multor țesuturi au proprietatea schimbării formei, dar această abilitate devine funcția principală în țesutul muscular.
Caracteristică generală și clasificare
Principalele semne morfologice ale elementelor țesutului muscular sunt forma alungită, prezența miofibrilelor și miofilamentelor localizate longitudinal - organele speciale care asigură contractilitatea, localizarea mitocondriilor în apropierea elementelor contractile, prezența incluziunilor glicogenului, lipidelor și mioglobinei.
Țesut muscular fin.
Această țesătură este formată din mezenchimă. Celulele musculare netede - miocite alungite cu capete fusiforme, acoperite cu o membrană celulară. Un miocit este o celulă mononucleară. Nucleul în formă de tijă ocupă o poziție centrală în celulă. Conține heterocromatină atomizată și unul sau doi nucleoli clar vizibili. Centrul celular (citocenter) este situat în apropierea uneia dintre suprafețele nucleului. Organelele rămase sunt concentrate la polii nucleului în formă de tijă. Mitocondrii bine dezvoltate, complex Golgi, EPS neted; ribozomi, granule EPS sunt ușor exprimate. Citoplasma miocitelor conține granule de glicogen - rezerva de energie a celulei.
Miocitele sunt strâns adiacente una de cealaltă, formând straturi și grupuri, separate de un țesut conjunctiv neformat. Miofibrilele sunt localizate în citoplasmă, merg de-a lungul periferiei celulei de-a lungul axei sale. Ele constau din fire subțiri și sunt un element contractil al mușchiului.
În exterior, fibra musculară este acoperită cu o membrană - un sarcolemă, format din straturile interioare și exterioare. Stratul interior este un plasmolemă, care este similar cu membrana altor celule tisulare. Stratul de țesut conjunctiv exterior este format dintr-o membrană a subsolului și structuri fibroase adiacente. Plasmolemma formează un sistem de tuburi înguste care pătrund în fibra musculară.
Fibrele de țesut conjunctiv situate în afara membranei subsolului din fibra musculară formează un endomiziu, care este bogat în vase de sânge și nervi. Endomysium se combină cu perimisium, o membrană care acoperă un grup de fibre musculare. Perimisiumul mai multor fascicule musculare este conectat cu epimisiul - membrana țesutului conjunctiv cel mai extern care combină mai multe dintre aceste mănunchiuri într-un mușchi - un organ caracterizat printr-o structură și funcție specifice.
Țesutul muscular neted se găsește în pereții stomacului, intestinelor, uterului, vezică urinară, uretere, bronhii, precum și în vase de sânge medii și mari. Activitatea musculară netedă este reglementată de ANS. Contracțiile musculare nu se supun voinței unei persoane și, prin urmare, țesutul muscular neted se numește musculatură involuntară.
Țesut muscular muscular striat.
Țesutul muscular este scheletul contractil al trunchiului, capului, membrelor, faringelui, laringelui, jumătății superioare a esofagului, limbii și mușchilor masticatori.
Se formează din miotomii somiților mezodermului. Unitatea structurală este fibra musculară striată. Are un corp cilindric, este acoperit cu o cochilie - sarcolem, iar citoplasma se numește - sarcoplasma, în care există numeroși nuclei și miofibrilele. Fiecare miofibril este format din discuri care au o compoziție chimică diferită. Miofibrilele sunt aparatul contractil al fibrei musculare.
Toți mușchii scheletului sunt construiți din țesutul muscular striat. Mușchii sunt arbitrari, deoarece contracția sa apare sub influența scoarței cerebrale.
Țesutul muscular al inimii.
Acest tip de țesut muscular formează membrana mijlocie a inimii - miocardul, prin natura reducerii se referă la involuntar, deoarece nu este controlat de voința animalului. Se dezvoltă din splanchnatoma frunzei viscerale - lamina miepicardică.
Țesutul muscular cardiac este format din celule musculare - cardiomiocite (miocite cardiace). Miocitele, care se conectează între ele la capetele lor de-a lungul axei lungi a celulelor, formează o structură similară cu fibra musculară.
Există două tipuri de celule. : celule contractile tipice și miocite cardiace atipiceconstituind sistemul de conducere a inimii.
Celule musculare tipice îndeplinește funcția contractilă; au formă dreptunghiulară, 1-2 nuclei sunt în centru, miofibrilele sunt situate la periferie. Între miocitele adiacente există discuri de inserție. Cu ajutorul lor, miocitele se colectează în fibrele musculare, separate printr-un țesut conjunctiv-fibros subțire. Între fibrele musculare adiacente se leagă fibre care asigură contracția miocardului în ansamblu.
Sistemul de conducere al inimii este format din fibre musculare constând din celule musculare atipice. Sunt mai mari decât contractile, mai bogate în sarcoplasmă, dar mai sărace decât miofibrilele, care se intersectează adesea. Sâmburii sunt mai mari și nu întotdeauna în centru. Fibrele sistemului conducător sunt înconjurate de un plex dens de fibre nervoase.
Țesuturile musculare (lat. Textus muscularis) se numesc țesuturi care sunt diferite ca structură și origine, dar similare în capacitatea de contracții pronunțate. Țesutul muscular este alcătuit din celule alungite din care iau iritația sistemul nervos și răspundeți cu o reducere. Ele asigură mișcarea în spațiul corpului în ansamblul său, mișcarea organelor sale în interiorul corpului (inimă, limbă, intestine etc.) și constă din fibre musculare. Celulele multor țesuturi au proprietatea schimbării formei, dar în țesuturile musculare această capacitate devine funcția principală.
Principalele semne morfologice ale elementelor țesutului muscular: o formă alungită, prezența miofibrilelor și miofilamentelor localizate longitudinal - organele speciale care asigură contractilitatea, localizarea mitocondriilor în apropierea elementelor contractile, prezența incluziunilor glicogenului, lipidelor și mioglobinei.
Organelele contractile speciale, miofilamentele sau miofibrilele asigură contracția care apare atunci când două proteine \u200b\u200bfibrilare principale - actină și miozină - interacționează cu ele, cu participarea obligatorie a ionilor de calciu. Mitocondriile furnizează energie acestor procese. Stocul surselor de energie este format din glicogen și lipide. Mioglobina este o proteină care asigură legarea oxigenului și crearea rezervei sale în momentul contracției musculare, când vasele de sânge sunt comprimate (aportul de oxigen scade brusc).
Proprietățile țesutului muscular:
- excitabilitate
- conductibilitate
- contractilitatea
- labilitate
Tipuri de țesut muscular:
1. Țesut muscular neted
Țesutul muscular neted este format din celule mononucleare - miocite în formă de fus cu o lungime de 20-500 microni. Citoplasma lor la un microscop ușor arată uniform, fără striație transversală. Acest țesut muscular are proprietăți speciale: se contractă lent și se relaxează, are automatizare, este involuntar (adică activitatea sa nu este controlată de voința unei persoane). Incluse în pereți organe interne: vase sanguine și limfatice, tractul urinar, tractul digestiv (contracția pereților stomacului și intestinelor).
2. Țesut muscular muscular schelet
Este format din miocite având o lungime mare (până la câțiva centimetri) și un diametru de 50-100 microni; aceste celule sunt multinucleate, conțin până la 100 sau mai mulți nuclei; într-un microscop ușor, citoplasma arată ca o alternanță de dungi întunecate și ușoare. Proprietățile acestui țesut muscular sunt o rată ridicată de contracție, relaxare și aleatoriu (adică activitatea sa este controlată de voința unei persoane). Acest țesut muscular face parte din mușchii scheletului, precum și pereții faringelui, partea superioară a esofagului, formează limba, mușchii oculomotori. Fibrele de la 10 la 12 cm lungime.
3. Țesut muscular muscular striat
Este format din 1 sau 2 cardiomiocite nucleare care au o striație transversală a citoplasmei (de-a lungul periferiei citolemmei). Cardiomiocitele sunt ramificate și formează conexiuni interconectate - discuri de inserție în care este combinat citoplasma lor Există, de asemenea, un alt contact intercelular - anostamosele (inhibarea citolemmei unei celule în citolema altei) Acest tip de țesut muscular formează miocardul inimii. Se dezvoltă din placa mioepicardică (frunza viscerală a splanchnotomului gâtului de embrion) O proprietate specială a acestui țesut este automatizarea - capacitatea de a se contracta și relaxa ritmic sub influența excitației care apare în celulele în sine (cardiomiocite tipice). Acest țesut este involuntar (cardiomiocite atipice). Există un al treilea tip de cardiomiocit - cardiomiocite secretoare (nu au fibrilele) sintetizează hormonul troponin, care scade tensiunea arterială și extinde pereții vaselor de sânge.
Membrana musculară a inimii (miocard) - este formată din celule ale cavității musculare, care sunt conectate la complexe musculare sau fibre musculare cardiace cu ajutorul discurilor de inserție. Țesutul muscular are capacitatea de a se reface, este protejat de țesut conjunctiv, formând o cicatrice.
Vă trimiteți munca bună la baza de cunoștințe. Folosiți formularul de mai jos
Studenții, studenții absolvenți, tinerii oameni de știință care folosesc baza de cunoștințe în studiile și munca lor vă vor fi foarte recunoscători.
Postat pe http://www.allbest.ru/
Țesut muscular combină capacitatea de a reduce.
Caracteristici structurale: aparatul contractil, care ocupă o parte semnificativă în citoplasma elementelor structurale ale țesutului muscular și este format din filamente de actină și miozină, care formează organele cu scop special - myofibrils .
Țesut muscularele sunt un grup de țesuturi de origine și structură variate, unite pe baza unei trăsături comune - o abilitate contractilă pronunțată, datorită căreia își pot îndeplini funcția principală - de a muta corpul sau părțile sale în spațiu.
Cele mai importante proprietăți ale țesutului muscular.Elementele structurale ale țesutului muscular (celule, fibre) au o formă alungită și sunt capabile să se contracte datorită dezvoltării puternice a aparatului contractil. Acesta din urmă se caracterizează printr-un aranjament extrem de ordonat actinași miofilamente de miozină,crearea condițiilor optime pentru interacțiunea lor. Acest lucru este realizat prin conectarea structurilor contractile cu elemente speciale ale citoscheletului și plasmolemmei (Sarcolemă)efectuarea funcția de referință. În partea țesutului muscular, miofilamentele formează organele cu semnificație specială - myofibrils.Pentru contracția musculară, este necesară o cantitate semnificativă de energie, prin urmare, în elementele structurale ale țesuturilor musculare există un număr mare de mitocondrii și incluziuni trofice (picături de lipide, granule de glicogen) care conțin substraturi - surse de energie. Deoarece contracția musculară are loc cu participarea ionilor de calciu, structurile care realizează acumularea și secreția acestuia sunt bine dezvoltate în celulele și fibrele musculare - reticulul agranular endoplasmic (reticul sarcoplasmic), caveola.
Clasificarea țesuturilor muscularepe baza semnelor structurii și funcției lor (a) (clasificare morfofuncțională)și (b) originea (clasificare histogenetică).
Clasificarea morfofuncțională a țesutului muscular scoate în evidență țesut muscular muscular striat (striat)și țesut muscular neted.Țesutul muscular striat este format din elemente structurale (celule, fibre), care au o striație transversală datorită dispunerii speciale ordonate reciproce a actinei și miofilamentelor de miozină din ele. Țesuturile musculare striate includ scheleticși tesutul muscular cardiac.Țesutul muscular neted este format din celule care nu au striație transversală. Cel mai frecvent tip de țesut este țesutul muscular neted, care face parte din peretele diferitelor organe (bronhii, stomac, intestine, uter, trompă uterină, ureter, vezică și vase de sânge).
Clasificarea histogenetică a țesutului muscular distinge trei tipuri principale de țesut muscular: somatic(țesut muscular muscular) coelomic(țesut muscular muscular) și mezenchimale(țesut muscular neted al organelor interne), precum și două suplimentare: celule mioepiteliale(celule contractile epiteliale modificate în secțiunile terminale și mici conducte excretoare ale unor glande) și elemente minerale(celule contractile de origine neurală în irisul ochiului).
Țesut muscular muscular striat (striat)în masa sa depășește orice alt țesut corporal și este cel mai frecvent țesut muscular al corpului uman. Oferă mișcarea corpului și a părților sale în spațiu și menține postura (o parte a aparatului locomotor), formează mușchi oculomotori, mușchii peretelui cavității bucale, limbă, faringe și laringe. Țesutul muscular striat visceral non-scheletic, care se găsește în treimea superioară a esofagului, este o parte a sfincterelor anale și uretrale externe.
miocit scheletic muscular
Țesutul muscular striat scheletal se dezvoltă în perioada embrionară din myotomessomiti care dau naștere la divizare activă mioblaste- celule care sunt dispuse în lanțuri și se contopesc între ele în regiunea capetelor cu formarea tuburi musculare (miotubule)transformându-se în fibre musculare.Astfel de structuri formate dintr-o singură citoplasmă uriașă și numeroși nuclei sunt menționate în mod tradițional în literatura rusă ca symplasts(în acest caz - myosymplasts)cu toate acestea, acest termen este absent în terminologia internațională acceptată. Unele mioblaste nu se contopesc cu altele, localizate pe suprafața fibrelor și dând naștere la miosatellitotsitam- celule mici, care sunt elemente cambiale ale țesutului muscular scheletic. Țesutul muscular scheletic format în mănunchiuri fibre musculare striate, care sunt unitățile sale structurale și funcționale.
Fibrele musculare țesutul muscular scheletic este formațiuni cilindrice de lungime variabilă (de la milimetri la 10-30 cm). De asemenea, diametrul lor variază foarte mult în funcție de apartenența la un anumit mușchi și tip, starea funcțională, gradul de încărcare funcțională, statutul nutrițional și alți factori. În mușchi, fibrele musculare formează mănunchiuri în care se află în paralel și, deformându-se reciproc, dobândesc adesea o formă multifacetă neregulată, care se vede mai ales în secțiunile transversale. Între fibrele musculare sunt straturi subțiri de fibroase libere țesut conjunctivtransportarea vaselor și nervilor - endomisium.Striația transversă a fibrelor musculare scheletice datorită alternării întunericului discuri anizotrope (benzile A)și luminos discuri izotrope (benziI). Fiecare disc izotrop este tăiat la jumătate de o întuneric subțire linia Z - telefragm. Nucleii fibrei musculare - relativ ușoare, cu 1-2 nucleoli, diploide, ovale, aplatizate - se află pe periferia acesteia sub sarcolemă și sunt localizate de-a lungul fibrei. În afara sarcolemmei este acoperită în gros membrana subsoluluiîn care sunt țesute fibrele reticulare.
Celulele miosatelitare (celule miosatelitare) - mici celule aplatizate localizate cu impresii superficiale ale sarcolemmei fibrei musculare și acoperite cu o membrană comună a subsolului (vezi Fig. 88). Nucleul miosatelitului este dens, relativ mare, organulele sunt mici și puține. Aceste celule sunt activate când fibrele musculare sunt deteriorate și asigură regenerarea lor reparativă. Fuzionându-se cu restul fibrei cu o sarcină crescută, celulele miosatelitare sunt implicate în hipertrofia sa.
myofibrils formează aparatul contractil al fibrei musculare, sunt localizate în sarcoplasmă de-a lungul lungimii sale, ocupând partea centrală și sunt evidențiate distinct pe secțiunile transversale ale fibrelor sub formă de puncte mici.
Miofibrilele au propria striație transversală și sunt atât de ordonate în fibra musculară încât discurile izotrope și anisotrope ale diferitelor miofibrilele coincid, provocând striația transversă a întregii fibre. Fiecare miofibril este format din mii de structuri repetate interconectate secvențial - sarcomeres.
inokomma (Myomere)este o unitate structurală și funcțională a miofibrilelor și reprezintă porțiunea sa situată între două teloperme (liniile Z).Include un disc anizotrop și două jumătăți de discuri izotrope - o jumătate pe fiecare parte. Sarcomere format dintr-un sistem ordonat gros (miozină)și miofilamente subțiri (active).Miofilamente groase sunt asociate cu mezofragm (linia M)și concentrat într-un disc anizotrop,
și miofilamente subțiri atașate la telograme (liniile Z),formează discuri izotrope și pătrund parțial în discul anizotrop, între filamente groase până la lumină banda Hîn centrul discului anisotrop.
La nivelul mușchilor, ca și în celelalte țesuturi, se disting două tipuri de regenerare - fiziologice și reparative. Regenerarea fiziologică se manifestă sub formă de hipertrofie a fibrelor musculare, care se exprimă printr-o creștere a grosimii și a lungimii lor uniforme, o creștere a numărului de organele, în principal miofibrilele și o creștere a numărului de nuclei, care se manifestă în cele din urmă într-o creștere a capacității funcționale a fibrelor musculare. Folosind metoda radioizotopului, s-a constatat că o creștere a numărului de nuclei din fibrele musculare în condiții de hipertrofie se realizează prin divizarea celulelor miosatelitare și intrarea ulterioară a celulelor fiice în miosimplast.
O creștere a numărului de miofibriluri se realizează prin sinteza proteinelor de actină și miozină prin ribozomi liberi și asamblarea ulterioară a acestor proteine \u200b\u200bîn miofilamente de actină și miozină în paralel cu filamentele de sarcomere corespunzătoare. Ca urmare a acestui fapt, la început are loc îngroșarea miofibrilelor, apoi clivajul lor și formarea miofibrilelor fiice. În plus, formarea de noi miofilamente de actină și miozină este posibilă nu în paralel, ci cu fundul miofibrilelor anterioare, realizând astfel alungirea lor. Reticulul sarcoplasmic și tubulii T din fibra hipertrofică se formează datorită proliferării elementelor precedente. În anumite tipuri antrenament muscular se poate forma un tip predominant roșu de fibre musculare (la stiers) sau un tip alb de fibre musculare (la sprinter). Hipertrofia legată de vârstă a fibrelor musculare se manifestă intens odată cu debutul activității motorii a corpului (1-2 ani), care se datorează în principal stimulării nervoase crescute. La vârsta senilă, precum și în condiții de încărcare musculară scăzută, are loc atrofierea organelelor speciale și generale, subțierea fibrelor musculare și scăderea capacității lor funcționale.
Regenerarea reparativă se dezvoltă după deteriorarea fibrelor musculare. Metoda de regenerare depinde de mărimea defectului. Cu o deteriorare semnificativă în întreaga fibră musculară, miosateliții din zona de deteriorare și din zonele adiacente sunt dezinhibați, proliferează intens și apoi migrează în zona defectului fibrei musculare, unde se aliniază pentru a forma un miotub. Diferențierea ulterioară a miotubului duce la completarea defectului și restabilirea integrității fibrei musculare. În condițiile unui mic defect al fibrei musculare la capetele sale, datorită regenerării organelelor intracelulare, se formează rinichi musculari care cresc unul față de celălalt și apoi se îmbină, ceea ce duce la închiderea defectului. Cu toate acestea, regenerarea reparativă și refacerea integrității fibrelor musculare poate fi efectuată în anumite condiții: în primul rând, cu inervarea motorie conservată a fibrelor musculare, iar în al doilea rând, dacă elementele de țesut conjunctiv (fibroblaste) nu intră în zona de deteriorare. În caz contrar, la locul defectului fibrei musculare se dezvoltă o cicatrice de țesut conjunctiv.
Savantul sovietic A.N. Studenții au dovedit posibilitatea autotransplantării țesutului muscular scheletal și chiar a mușchilor întregi în anumite condiții:
· Măcinarea mecanică a țesutului muscular al transplantului, cu scopul dezinhibării celulelor satelite și a proliferării ulterioare a acestora;
· Amplasarea țesutului mărunțit în patul fascial;
· Există fibra nervului motor la grefa zdrobită;
· Prezența mișcărilor contractile de antagoniști și sinergici musculari.
Din punct de vedere anatomic, nou-născuții au toți mușchii scheletici, dar în ceea ce privește greutatea corporală, acestea constituie doar 23% (la un adult 44%). Cantitatea de fibre musculare din mușchi este aceeași ca la un adult. Cu toate acestea, microstructura fibrelor musculare este diferită: fibrele au diametrul mai mic, au mai mulți nuclei. Pe măsură ce crește, există o îngroșare și o alungire a fibrelor. Acest lucru se datorează îngroșării miofibrilelor, împingând nucleul la periferie. Mărimea fibrelor musculare se stabilizează cu 20 de ani.
Mușchii la copii sunt mai elastici decât la adulți. Ie scurtați mai repede când vă contractați și prelungiți-vă când vă relaxați. Excitabilitatea și labilitatea mușchilor nou-născuților este mai mică decât cea a adulților, dar crește odată cu vârsta. La nou-născuți, chiar și în vis, mușchii sunt într-o stare de ton. Dezvoltarea diferitelor grupe musculare are loc inegal. La 4-5 ani, mușchii antebrațului sunt mai dezvoltați, mușchii mâinii rămân în urmă în dezvoltare. Încălzirea accelerată a mușchilor mâinii are loc în 6-7 ani. Mai mult, extensoarele se dezvoltă mai lent decât flexorii. Odată cu vârsta, raportul tonusului muscular se modifică. În copilărie timpurie, tonul crescut al mușchilor mâinii, extensorii șoldului etc. treptat se normalizează distribuția tonului.
Inima ca organ se caracterizează prin capacitatea de a se regenera prin hipertrofie regenerativă, în care masa organului este restabilită, dar forma rămâne perturbată. Un fenomen similar este observat după infarctul miocardic, când masa inimii se poate recupera în ansamblu, în timp ce la locul de deteriorare se formează o cicatrice de țesut conjunctiv, dar hipertrofiile de organe, adică. forma este ruptă. Nu numai o creștere a dimensiunii cardiomiocitelor apare, dar și proliferarea în principal în atrii și urechile inimii.
Se credea anterior că diferențierea cardiomiocitelor este un proces ireversibil asociat cu pierderea completă a capacității de divizare a celulelor. Dar la nivelul actual, numeroase date arată că cardiomiocitele diferențiate sunt capabile de sinteza ADN-ului și mitozei. În lucrările de cercetare ale P.P. Rumyantsev și studenții săi au arătat că, după infarctul miocardic ventricular stâng experimental, 60-70% din cardiomiocitele atriale revin la ciclul celular, numărul de celule poliploide crește, dar acest lucru nu compensează leziunile miocardului.
S-a constatat că cardiomiocitele sunt capabile de diviziune mitotică (inclusiv celulele sistemului de conducere). În miocardul inimii, există în special multe celule poliploide mononucleare cu un conținut de ADN de 16-32 de ori, dar există și cardiomiocite binucleare (13-14%), în principal octoploid.
În procesul de regenerare a țesutului muscular cardiac, cardiomiocitele participă la procesul de hiperplazie și hipertrofie, ploidia lor crește, dar nivelul de proliferare a celulelor țesutului conjunctiv în zona de deteriorare este de 20-40 de ori mai mare. În fibroblaste, sinteza de colagen este activată, ca urmare a unei reparații prin cicatrizarea defectului. Prezentarea biologică a unei astfel de reacții adaptive a țesutului conjunctiv este explicată prin importanța vitală a organului cardiac, deoarece întârzierea închiderii defectului poate duce la moarte.
Se credea că la nou-născuți și, probabil, la copilăria timpurie, când sunt capabili să divizeze cardiomiocitele încă persistă, procesele regenerative sunt însoțite de o creștere a numărului de cardiomiocite. Mai mult, la adulți, regenerarea fiziologică se realizează în miocard în principal prin regenerare intracelulară, fără a crește numărul de celule, adică. în miocardul unui adult, nu există proliferare de cardiomiocite. Însă recent s-au obținut dovezi că într-o inimă umană sănătoasă, 14 dintr-un milion de miocite se află într-o stare de mitoză, rezultând o citotomie, adică. numărul de celule nu este semnificativ, dar în creștere.
Utilizarea metodelor moderne de biologie celulară în studii clinice și experimentale ne-a permis să continuăm să elucidăm mecanismele celulare și moleculare de deteriorare și regenerare miocardică. Un interes deosebit este dovada că în regiunile perinecrotice și într-o inimă congestionată funcțional are loc sinteza proteinelor și peptidelor miocardice embrionare, precum și a proteinelor sintetizate în timpul ciclului celular. Acest lucru confirmă similaritatea dintre mecanismele de regenerare și ontogeneza normală.
De asemenea, s-a dovedit că cardiomiocitele diferențiate în cultură sunt capabile de diviziune mitotică activă, ceea ce poate fi explicat nu prin pierderea completă, ci printr-o suprimare a capacității cardiomiocitelor de a reveni la ciclul celular.
O sarcină importantă a cardiologiei teoretice și practice este dezvoltarea unor modalități de stimulare a restaurării miocardului deteriorat, adică. inducerea regenerării miocardului și reducerea cicatricii țesutului conjunctiv. Unul dintre domeniile de cercetare oferă posibilitatea transferului genelor regulatoare care transformă fibroblastele cicatrice în mioblaste sau transfecția în cardiomiocite ale genelor care controlează creșterea celulelor noi. O altă direcție este transferul către zona de afectare a celulelor scheletului și miocardului fetal, care ar putea fi implicate în refacerea mușchiului cardiac. Există, de asemenea, experimente privind transplantul de mușchi scheletici în inimă, care arată formarea zonelor de contractare a țesutului în miocard și îmbunătățirea indicilor funcționali miocardici. Tratamentul cu utilizarea factorilor de creștere care au efecte directe și indirecte asupra miocardului deteriorat, de exemplu, îmbunătățirea angiogenezei, poate fi promițător.
Țesut muscular fin
Trei grupuri de țesuturi musculare netede (sau nestriate) se disting prin origine - mezenchimale, epidermice și neurale.
Țesut muscular de origine mezenchimală
Histogeneză. Celulele stem și celulele progenitoare ale țesutului muscular neted, fiind deja determinate, migrează în locurile organelor ouătoare. Diferențindu-se, acestea sintetizează componentele matricei și colagenul membranei subsolului, precum și elastina. În celulele definitive (miocite), capacitatea sintetică este redusă, dar nu dispare complet.
Unitatea structurală și funcțională a țesutului muscular neted sau netratat este o celulă musculară netedă, sau un miocit neted este o celulă în formă de fus 20-500 μm lungime, 5-8 μm lățime. Nucleul celulei este în formă de tijă, situat în partea sa centrală. Când miocitul se contractă, miezul său se îndoaie și chiar se răsucește. Organele de importanță generală, printre care se află multe mitocondrii, sunt concentrate în citoplasmă în apropierea poliilor nucleului. Aparatul Golgi și reticulul endoplasmic granular sunt slab dezvoltate, ceea ce indică o activitate scăzută a funcțiilor sintetice. Ribozomii sunt localizați în mare parte liber.
Filamentele de actină formează o rețea tridimensională în citoplasmă, alungită în principal longitudinal sau mai degrabă oblic-longitudinal. Capetele filamentelor sunt legate între ele și plasmolemmei cu proteine \u200b\u200breticulante speciale. Aceste zone sunt clar vizibile în micrografele electronice sub formă de corpuri dense.
Filamentele de miozină sunt într-o stare depolimerizată. Monomerii de miozină sunt localizați lângă filamentele de actină. Semnalul contracției vine de obicei prin fibrele nervoase. Mediatorul, care iese în evidență din terminalele lor, schimbă starea plasmolemmei. Formează intruziuni - caveola, în care sunt concentrați ionii de calciu. Caveolae este așezată spre citoplasmă sub formă de bule (aici calciul este eliberat din bule). Aceasta implică atât polimerizarea miozinei, cât și interacțiunea miozinei cu actina. Filamentele de actină sunt deplasate unul spre celălalt, petele dense se reunesc, forța este transmisă membranei plasmatice și întreaga celulă este scurtată. Când semnalele sistemului nervos încetează, ionii de calciu sunt evacuați din caveole, miozina este depolimerizată și miofibrilele se degradează. Astfel, complexele actină-miozină există în miocite netede numai în perioada de contracție.
Miocitele netede sunt localizate fără spații intercelulare vizibile și sunt separate de o membrană a subsolului. În anumite zone, în el se formează „ferestre”, astfel încât plasmolemma miocitelor vecine converg. Aici, se formează nexuzele și nu numai mecanice, ci și legături metabolice apar între celule. În partea de sus a „copertelor” de la membrana subsolului dintre miocite sunt fibre elastice și reticulare, care combină celulele într-un singur complex tisular. Fibrele reticulare pătrund în fisurile de la capetele miocitelor, se fixează acolo și transmit forța contracției celulare întregii lor asociații.
Regenerarea. Regenerarea fiziologică a țesutului muscular neted se manifestă în condiții de încărcare funcțională crescută. Acest lucru se vede cel mai clar în membrana musculară a uterului în timpul sarcinii. O astfel de regenerare se realizează nu atât la nivelul țesutului, cât și la nivel celular: miocitele cresc, procesele sintetice sunt activate în citoplasmă, numărul de miofilamente crește (hipertrofia celulelor care lucrează). Cu toate acestea, proliferarea celulară (adică hiperplazia) nu este exclusă.
Ca parte a organelor, miocitele sunt combinate în mănunchiuri, între care există straturi subțiri de țesut conjunctiv. Fibrele reticulare și elastice din jurul miocitelor sunt țesute în aceste straturi. Vasele de sânge și fibrele nervoase trec prin straturile intermediare. Terminalele acestora din urmă nu se termină direct pe miocite, ci între ele. Prin urmare, după sosirea unui impuls nervos, mediatorul se răspândește difuz, excitând multe celule simultan. Țesutul muscular neted de origine mezenchimală este reprezentat mai ales în pereții vaselor de sânge și în multe organe interne tubulare și formează, de asemenea, mușchi mici individuali.
Țesutul muscular neted în anumite organe are proprietăți funcționale inegale. Acest lucru se datorează faptului că pe suprafața organelor există receptori diferiți pentru substanțe biologice specifice biologic. Prin urmare, pentru multe medicamente, reacția lor nu este aceeași.
Țesut muscular fin de origine epidermică
Celulele mioepiteliale se dezvoltă din germenul epidermic. Se găsesc în glande sudoripare, mamare, salivare și lacrimale și au precursori comuni cu celulele secretoare glandulare. Celulele mioepiteliale sunt atașate direct de epiteliu corespunzător și au o membrană comună a subsolului cu ele. În timpul regenerării, celulele și alte celule sunt restaurate din precursorii obișnuiți cu diferențe reduse. Majoritatea celulelor mioepiteliale au formă de stea. Aceste celule sunt adesea numite în formă de coș: procesele lor acoperă secțiunile de capăt și conductele mici ale glandelor. Nucleul și organulele de importanță generală sunt localizate în corpul celulei, iar aparatele contractile, organizate, ca și în celulele țesutului muscular de tip mezenchimal, sunt localizate în procese.
Țesut muscular neted de origine neurală
Miocitele acestui țesut se dezvoltă din celulele rudimentului neural ca parte a peretelui intern al gobletului optic. Corpurile acestor celule sunt situate în epiteliul suprafeței posterioare a irisului. Fiecare dintre ele are un proces care merge în grosimea irisului și se află paralel cu suprafața sa. În proces există un aparat contractil, organizat în același mod ca în toate miocitele netede. În funcție de direcția proceselor (perpendicular sau paralel cu marginea pupilei), miocitele formează doi mușchi - o pupilă care se îngustează și se extinde.
concluzie
După cum s-a menționat deja, țesutul muscular este un grup de țesuturi corporale de origine diversă, unite prin principiul contractilității: striate (scheletice și cardiace), netede, precum și țesuturi contractile specializate - epiteliale-musculare și neurogliale, care face parte din irisul ochiului.
Țesutul mușchiului scheletal striat apare din miotomii care alcătuiesc elementele mezodermului segmentat - somite.
Țesutul muscular neted al omului și vertebratelor se dezvoltă ca parte a derivaților mezenchimului, precum și a țesutului intern. Cu toate acestea, toate țesuturile musculare se caracterizează printr-o izolare similară în embrionul embrionar sub formă de celule în formă de fus - celule formatoare de mușchi sau mioblaste.
Contracția fibrelor musculare implică scurtarea miofibrilelor în fiecare sarcomere. Filamente groase (miozină) și subțiri (actină), într-o stare relaxată, conectate numai de secțiunile de capăt, în momentul contracției, efectuează mișcări de alunecare unul spre celălalt. Eliberarea de energie necesară reducerii energiei are loc ca urmare a transformării ATP în ADP sub influența miozinei. Activitatea enzimatică a miozinei se manifestă în condiția conținutului optim de Ca2 +, care se acumulează în reticulul sarcoplasmic.
Referințe
1. Histologie. Editat de Yu.I. Afanasyeva, N.A. Yurina. M .: „Medicină”, 1999
2. R. Eckert, D. Rendel, J. Augustine „Fiziologia animalelor” - 1 vol. M.: „Mir”, 1981
3. K.P. Ryabov „Istologia cu elementele de bază ale embriologiei” Minsk: „Școala superioară”, 1990
4. Histologie. Editat de Ulumbekov, prof. Univ. YA Chelisheva. M .: 1998
5. Histologie. Editat de V.G. Eliseev. M .: „Medicină”, 1983
Postat pe Allbest.ru
...Documente similare
Caracteristicile structurale ale țesutului muscular. Studiul mecanismului de contracție musculară și a aparatului de transmitere a excitației. Histogeneza și regenerarea țesutului muscular. Principiile cardiomiocitelor contractile, conductoare și secretoare ale țesutului muscular al inimii.
cheat sheet adăugat la 11/11/2010
Studiul tipurilor și funcțiilor diferitelor țesuturi umane. Sarcini ale științei histologiei, care studiază structura țesuturilor organismelor vii. Caracteristici structurale ale țesutului epitelial, nervos, muscular și ale țesuturilor mediului intern (conjunctiv, scheletic și fluid).
prezentare adăugată la 11.08.2013
Studiul caracteristicilor structurale ale țesuturilor animale, funcționării și soiurilor. Efectuarea unui studiu privind o trăsătură caracteristică a structurii țesutului conjunctiv și nervos. Structura epiteliului plat, cubic, ciliat și glandular. Tipuri de țesut muscular.
prezentare adăugată la 02/08/2015
Caracteristici generale și caracteristici legate de vârstă ale țesutului cartilaj. Tipuri de cartilaj și țesut osos. Caracteristici generale și caracteristici legate de vârstă ale țesutului osos. Caracteristici ale structurii țesutului muscular în copilărie și bătrânețe. Țesut muscular muscular.
prezentare, adăugat 02/07/2016
Clasificarea țesuturilor, tipurile de țesuturi epiteliale, structura și funcțiile acestora. Funcția de susținere, trofică și protectoare a țesuturilor conjunctive. Funcțiile țesuturilor nervoase și musculare. Conceptul de organe și sisteme de organe, diferențele lor individuale, de sex, de vârstă.
rezumat, adăugat 09/11/2009
Țesutul epitelial, capacitatea sa de regenerare. Țesuturi conective implicate în menținerea homeostaziei mediului intern. Celule de adăpost și limfă. Țesut muscular muscular striat și cardiac. Funcțiile celulelor nervoase și țesuturile organismelor animale.
rezumat, adăugat 16 ianuarie 2015
Studiul tipurilor de țesut animal, precum și funcțiile pe care le îndeplinesc. Caracteristici structurale ale grupurilor de țesuturi epiteliale, conjunctive, musculare și nervoase. Determinarea locației fiecărui grup și importanța pentru viața animalului.
prezentare, adăugată 18/10/2013
Conceptul general și tipurile de vibrații. Caracterizarea proceselor de tensiune (compresie), forfecare, îndoire, torsiune. Proprietățile mecanice ale țesutului osos și vascular. Specificitatea țesutului muscular, principalele moduri de lucru muscular - izometric și izotonic.
lucrare de control, adăugată 19.03.2014
Celula ca principală unitate structurală a corpului. Descrierea structurii sale, a proprietăților vitale și chimice. Structura și funcțiile țesutului epitelial și conjunctiv, muscular și nervos. Organe și o listă a sistemelor de organe umane, scopul și funcțiile lor.
prezentare adăugată la 19.04.2012
Fiziologia și biochimia activității musculare ca o componentă importantă a metabolismului în organism. Tipuri de țesut muscular și, în consecință, mușchi care diferă în structura fibrelor musculare, natura inervației. Efectul activității fizice cu intensitate diferită.
Capitolul 9. TESTE MUSCULARE
Capitolul 9. TESTE MUSCULARE
Țesut muscular (textus muscularis)numite țesături care sunt diferite ca structură și origine, dar similare în capacitatea de a reduce. Ele asigură mișcarea în spațiul corpului în ansamblul său, părțile sale și mișcarea organelor în interiorul corpului (inimă, limbă, intestine etc.).
Celulele multor țesuturi au proprietatea de a se contracta cu o schimbare de formă, dar în țesuturile musculare această capacitate devine funcția principală.
9.1. CARACTERISTICI GENERALE MORFOFUNCȚIONALE ȘI CLASIFICARE
Principalele semne morfologice ale elementelor țesutului muscular sunt forma alungită, prezența miofibrilelor și miofilamentelor localizate longitudinal - organele speciale care asigură contractilitatea, localizarea mitocondriilor în apropierea elementelor contractile, prezența incluziunilor glicogenului, lipidelor și mioglobinei.
Organele contractile speciale - miofilamente sau miofibrilele asigură contracția care apare atunci când două proteine \u200b\u200bfibrilare principale - actină și miozină - interacționează cu ele, cu participarea obligatorie a ionilor de calciu. Mitocondriile furnizează energie acestor procese. Stocul surselor de energie este format din glicogen și lipide. Mioglobina este o proteină care asigură legarea oxigenului și crearea rezervei sale în momentul contracției musculare, când vasele de sânge sunt comprimate (fluxul de oxigen scade brusc).
Clasificare.Clasificarea țesutului muscular se bazează pe două principii - morfofuncționale și histogenetice. În conformitate cu principiul morfologic și funcțional, în funcție de structura organelelor de contracție, țesutul muscular este împărțit în două subgrupuri.
Primul subgrup- tesut muscular striat (striat) (textus muscularis striatus).În citoplasma elementelor lor, miosină filamen-
sunteți constant polimerizați, formați miofibrilele existente în mod constant cu filamente de actină. Acestea din urmă sunt organizate în complexe caracteristice - sarcomeres.În miofibrilele vecine, subunitățile structurale ale sarcomerelor sunt situate la același nivel și creează striația încrucișată.
Al doilea subgrup- țesut muscular neted (nelimitat) (textus muscularis nonstriatus).Aceste țesuturi se caracterizează prin faptul că, fără contracție, filamentele de miozină sunt depolimerizate. În prezența ionilor de calciu, acestea polimerizează și interacționează cu filamentele de actină. Miofibrilele formate în acest caz nu au o striație transversală: cu culori speciale sunt reprezentate de filamente (netede) uniform colorate pe întreaga lungime.
În conformitate cu principiul histogenetic, în funcție de sursele de dezvoltare (primordie embrionară), țesuturile musculare și elementele musculare sunt împărțite în: somatic (miotom), coelomic (din placa mioepicardică a foii de splanchnotom visceral), mezenchimal (din germenul desmal din mezenchimă (neural), neural tub neural), epidermic (din ectodermul cutanat și din placa prechordală).
9.2. ȚESURI MUSCULARE STRIPPATE
Există două varietăți principale de țesuturi striate (striate) - scheletul (miotomul) și cel cardiac (coelomic).
9.2.1. Țesut muscular muscular
Histogeneză.Sursa de dezvoltare a elementelor țesutului muscular striat scheletic (somatic) (textus muscularis striatus sceletalis)sunt celule stem ale miotomilor - promioblaste. Unii dintre ei se diferențiază la fața locului și participă la formarea așa-numitelor mușchi autohtoni. Alte celule migrează de la miotomi către mezenchimă. Acestea sunt deja determinate, deși în exterior nu diferă de alte celule mezenchimice. Diferențierea lor continuă la locurile unde sunt așezați alți mușchi ai corpului. În timpul diferențierii, apar două linii celulare. Celulele uneia dintre linii se contopesc, formând simplaste alungite - tuburi musculare (miotuburi). În ele, diferențierea organelelor speciale - miofibrilele (Fig. 9.1). În acest moment, în miotuburi se remarcă un reticul endoplasmic granular bine dezvoltat. Miofibrilele sunt mai întâi situate sub plasmolemă, apoi umplu majoritatea miotuburilor. Dimpotrivă, nucleele din departamentele centrale sunt mutate la periferie. Centrele celulare și microtubulele dispar complet. Endo granular
Fig. 9.1.Histogeneza țesutului muscular scheletic (conform A. A. Klishov):
și- promioblaste; b- miosimplast; în- tubul muscular; g- mușchi matur
fibre. 1 - myosatellite; 2 - miez de miosimplast; 3 - miofibrilele
reteaua de plasma este redusa semnificativ. Astfel de structuri definitive se numesc miosimplaste.
Celulele celeilalte linii rămân independente și se diferențiază în celulele miosatelitare. Aceste celule sunt localizate pe suprafața miosimplastelor. Celulele miosatelitare, care se înmulțesc, se contopesc cu miosimplaste, participând astfel la crearea unui nucleu optim
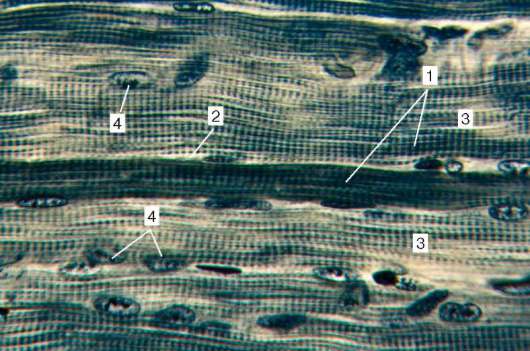
Fig. 9.2.Structura țesutului muscular striat (micrografie):
1 - fibre musculare; 2 - sarcolemă; 3 - sarcoplasma și miofibrilele; 4 - nuclee
myosymplast. Colorare - Hematoxilină de fier
relație sarcoplasmatică necesară pentru sinteza proteinelor specifice simplastului.
Structura.Unitatea structurală principală a țesutului muscular scheletic este fibra musculară, formată din miosimplast și miosatelocite, acoperite cu o membrană comună a subsolului (Fig. 9.2-9.4). Lungimea întregii fibre poate fi măsurată în centimetri cu o grosime de 50-100 microni. Complexul, format din plasmolemma miosimplastului și membrana subsolului, se numește sarcolemă.
Structura miosimplastului.Myosymplast are multe nuclee alungite situate direct sub plasmolemă. Numărul lor într-un simplast poate ajunge la câteva zeci de mii (vezi Fig. 9.2). Organele de importanță generală - complexul Golgi și fragmente mici din reticulul endoplasmatic agranular - sunt localizate la poli ai nucleelor. Miofibrilele umplu partea principală a miosimplastului și sunt amplasate longitudinal (vezi Fig. 9.3).
inokomma- unitatea structurală a miofibrilelor. Fiecare miofibril are discuri întunecate și luminoase transversale, cu refracție inegală (discuri A anizotrope și discuri I izotrope). Fiecare miofibril este înconjurat de bucle ale unui reticul endoplasmic agranular - un reticul sarcoplasmic - localizat longitudinal și anastomos între ele. Sarvenrele vecine au o structură de frontieră comună - linia Z (Fig. 9.5). Este construită sub forma unei rețele de molecule de fibrilă proteică, printre care alfa-actinina joacă un rol semnificativ. Capetele filamentelor de actină sunt conectate la această rețea. Din liniile Z vecine, filamentele de actină sunt direcționate spre centrul sarcomerei, dar nu ajung la mijlocul său. Filamente de actină combinate cu linia Z și filamentele de miozină
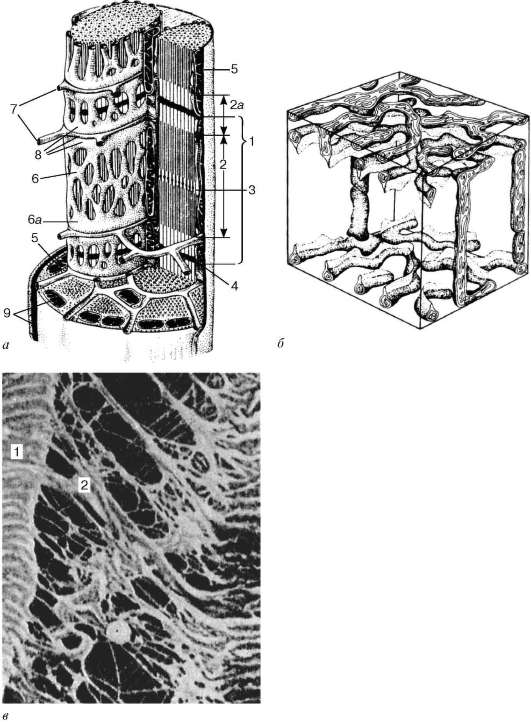
Fig. 9.3.Schema structurii ultramicroscopice a miosimplastului (conform R.V. Krstic, cu modificări) (a): 1 - sarcomere; 2 - disc anizotrop (banda A); 2a - disc izotrop (banda I); 3 - linia M (mezofragmă) în mijlocul discului anisotrop; 4 - linia Z (telofragmă) în mijlocul unui disc izotrop; 5 - mitocondrii; 6 - reticulul sar-coplasmic; 6a - rezervor final; 7 - tub transvers (tub T); 8 - triadă; 9 - sarcolemă; b- Dispunerea spațială a mitocondriilor în simplast. Planurile superioare și inferioare ale figurii sunt limitate de sarco-măsura discului anisotrop (conform L. E. Bakeeva, V. P. Skulachev, Yu. S. Chentsov); în- endomiziu. Micrografie electronică de scanare, mărire 2600 (preparat de Yu. A. Khoroshkov): 1 - fibre musculare; 2 - fibrilele de colagen
molecule de nebulină inextensibile fibrilare. În mijlocul discului întunecat al sarcomerei se află o rețea formată din miomiozină. Formează o linie M în secțiune transversală. La nodurile acestei linii M, capetele filamentelor de miozină sunt fixate. Celelalte capete ale acestora sunt îndreptate către liniile Z și
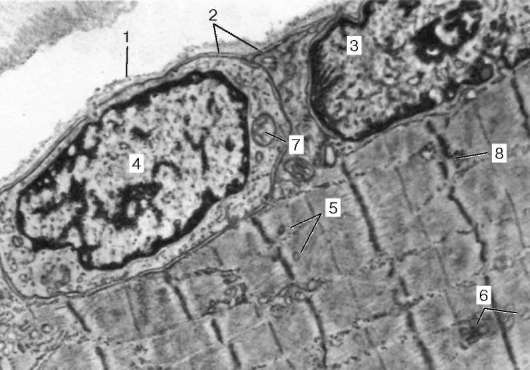
Fig. 9.4.Suprafața miosimplastului și a miosatelitului. Micrografie electronică, mărire 10.000 (preparat de V. L. Goryachkina și S. L. Kuznetsova): 1 - membrană subsol; 2 - plasmolemă; 3 - miezul miosimplastului; 4 - nucleul mios-tellitocitului; 5 - miofibrilele; 6 - tuburi ai rețelei endoplasmice (sarcoplasmatice) agranulare; 7 - mitocondrii; 8 - glicogen
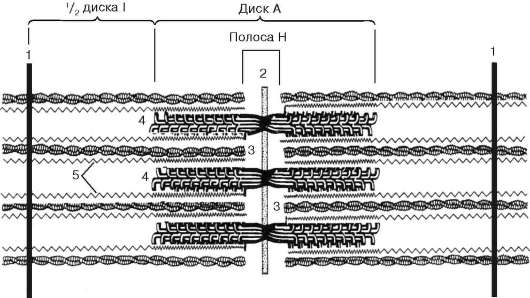
Fig. 9.5.Sarcomere (diagrama):
1 - linia Z; 2 - linia M; 3 - filamente de actină; 4 - filamente de miozină; 5 - molecule de titină fibrilară (conform B. Alberts, D. Bray, J. Lewis și colab., Modificate)
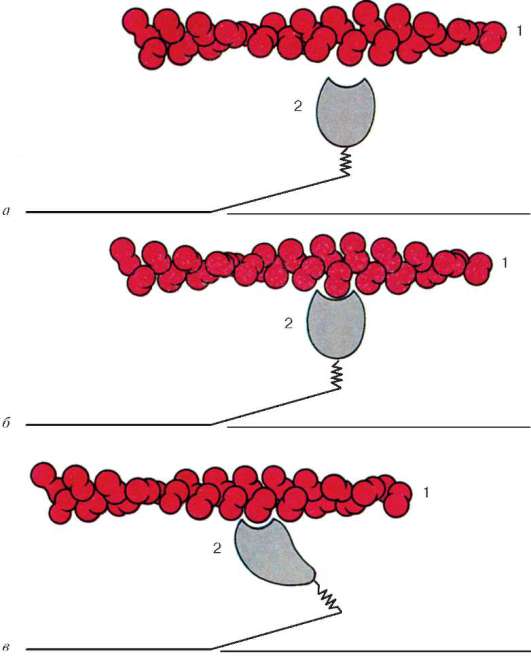
Fig. 9.6.Modificări conformaționale care implică o deplasare reciprocă a filamentelor de actină și miozină:
aC- modificări succesive în relațiile spațiale. 1 - actină; 2 - capul moleculei de miozină (conform B. Alberts, D. Bray, J. Lewis și colab., Modificat)
localizate între filamentele de actină, dar, de asemenea, nu ajung la liniile Z în sine. În același timp, aceste capete sunt fixate în raport cu liniile Z de către moleculele proteice gigantice de tracțiune din titină.
Moleculele de miozină au o coadă lungă și două capete la unul dintre capetele sale. Cu o creștere a concentrației ionilor de calciu în zona de atașare a capetelor (porțiunea balamalei), molecula își schimbă configurația (Fig. 9.6). În același timp (întrucât actina situată între filamentele de miozină), capetele de miozină se leagă de actină (cu participarea proteinelor auxiliare - tropomiozină și troponină). Apoi capul de miozină se înclină și trage molecula de actină împreună cu acesta spre linia M. Liniile Z converg, sarcomerul este scurtat.
Rețelele alfa-actinină ale liniilor Z ale miofibrilelor vecine sunt interconectate prin filamente intermediare. Ele se apropie de suprafața interioară a plasmolemmei și sunt fixate în stratul său cortical, astfel încât sarcomerele tuturor miofibrilelor sunt localizate la același nivel. Acest lucru, atunci când este observat la un microscop, creează impresia striației transversale a întregii fibre.
Sursa ionilor de calciu sunt rezervoarele reticulului endoplasmic agranular. Sunt alungite de-a lungul miofibrilelor de lângă fiecare sarcom și formează un reticul sarcoplasmic. În ea se acumulează ioni de calciu atunci când miosimplastul este într-o stare relaxată. La nivelul liniilor Z (în amfibieni) sau la granița discurilor A și I (la mamifere), tubulele rețelei se schimbă în direcție și sunt transversale, formând rezervoare extinse sau laterale (L).
De la suprafață până în adâncimea miosimplastului, plasmolemma formează tubuli lungi care se extind transversal (tuburi T) la nivelul limitelor dintre discurile întunecate și luminoase. Când miosimplastul primește un semnal despre începutul contracției, acesta se deplasează de-a lungul plasmolemmei sub formă de potențial de acțiune și se răspândește la membrana tubului T. Deoarece această membrană este aproape de membranele reticulului sarcoplasmic, starea acesteia din urmă se schimbă, calciul este eliberat din rezervoarele rețelei și interacționează cu complexele actină-miozină (acestea se contractă). Când potențialul de acțiune dispare, calciul se acumulează din nou în tubulele rețelei și contracția miofibrilelor se oprește. Energia este necesară pentru a dezvolta un efort de reducere. Este eliberat prin conversia ATP în ADP. Rolul ATPazei este îndeplinit de miozină. Sursa ATP este în principal mitocondrii, motiv pentru care sunt situate direct între miofibrilele.
Un rol important în activitatea miosimplastelor îl joacă incluziunile mioglobinei și glicogenului. Glicogenul servește ca sursă de energie, necesară nu numai pentru efectuarea muncii musculare, ci și pentru menținerea echilibrului termic al întregului organism. Mioglobina leagă oxigenul atunci când mușchiul este relaxat și sângele curge liber prin vasele mici de sânge. În timpul contracției musculare, vasele de sânge sunt comprimate, iar oxigenul depozitat este eliberat și este implicat în reacții biochimice.
Celulele miosatelitare.Aceste celule slab diferențiate sunt o sursă de regenerare a țesuturilor musculare. Ele sunt adiacente suprafeței miosimplastului, astfel încât plasmolemele lor sunt în contact (vezi fig. 9.1, 9.4). Celulele miosatelitare sunt mononucleare, nucleele lor întunecate sunt ovale și mai mici decât în \u200b\u200bsimplaste. Ei posedă toate organele de importanță generală (inclusiv centrul celular).
Tipuri de fibre musculare.Diferiți mușchi (precum organele) funcționează în diferite condiții biomecanice. Prin urmare, fibrele musculare din compoziția diferiților mușchi au rezistență, viteză și durată de contracție diferite, precum și oboseală. Activitatea enzimelor din ele este diferită și sunt prezentate sub diferite forme izomerice. Conținutul enzimelor respiratorii - glicolitice și oxidative - este, de asemenea, diferit.
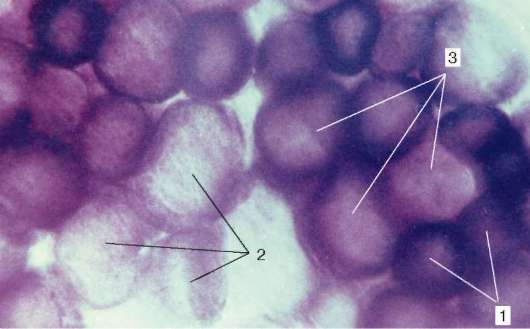
Fig. 9.7.Activitatea succinat dehidrogenazei în fibrele musculare de diferite tipuri (medicament V. F. Chetvergova, prelucrare conform Nakhlas și colab.): 1 - ridicat; 2 - scăzut; 3 - mediu
Prin raportul miofibrilelor, mitocondriilor și mioglobinei, se disting fibrele albe, roșii și intermediare. După caracteristicile funcționale, fibrele musculare sunt împărțite în rapid, lent și intermediar, care este determinat de organizarea moleculară a miozinei. Printre izoformele sale, există două principale - „rapid” și „lent”. Când se efectuează reacții histochimice, acestea sunt identificate prin activitatea ATPazei. Activitatea enzimelor respiratorii se corelează, de asemenea, cu aceste proprietăți. De obicei, procesele glicolitice predomină în fibre rapide, sunt bogate în glicogen, au mai puțin mioglobină, de aceea sunt numite albe. În fibrele lente, dimpotrivă, activitatea de enzime oxidante este mai mare, sunt mai bogate în mioglobină și arată mai roșu.
Alături de alb și roșu, există fibre intermediare. În majoritatea mușchilor scheletici, fibrele de diferite tipuri histochimice sunt mozaic (Fig. 9.7).
Proprietățile fibrelor musculare se schimbă odată cu schimbarea încărcărilor - sportive, profesionale, precum și în condiții extreme (greutate). Când reveniți la activități normale, astfel de modificări sunt reversibile. În unele boli (atrofia musculară, distrofia, consecințele denervării), fibrele musculare cu proprietăți inițiale diferite se schimbă diferit. Acest lucru vă permite să clarificați diagnosticul, pentru care sunt examinate exemplare de biopsie ale mușchilor scheletici.
Regenerarea.Nucleii miosimplastelor nu se pot diviza, deoarece nu există centre celulare în sarcoplasmă. Elementele cambiale servesc celulele miosatelitare.În timp ce corpul crește, acestea se divid, iar celulele fiice se contopesc cu miosimplaste. La sfârșitul creșterii, reproducerea celulelor miosatelitare se estompează. După deteriorarea fibrei musculare pentru o anumită măsură de la locul accidentării, aceasta este distrusă și fragmentul acesteia
ești fagocitată de macrofage. Restaurarea țesuturilor se realizează prin două mecanisme: hipertrofia compensatorie a simplastului propriu-zis și proliferarea celulelor miosatelitare. În simplast, se activează reticulul endoplasmic granular și complexul Golgi. Sinteza substanțelor necesare pentru restaurarea sarcoplasmei și a miofibrilelor are loc, precum și asamblarea membranelor, astfel încât integritatea limei plasmatice să fie restabilită. În acest caz, capătul deteriorat al miosimplastului se îngroașă, formând un rinichi muscular. Celulele miosatelitare care supraviețuiesc în apropierea leziunii sunt împărțite. Unii dintre ei migrează spre rinichiul muscular și se integrează în acesta, în timp ce alții se contopesc (la fel ca mioblastele în timpul histogenezei) și formează miotuburi noi care se dezvoltă în fibrele musculare.
9.2.2. Mușchiul scheletului ca organ
Transmiterea eforturilor de contracție către schelet se face prin tendoane sau prin atașarea mușchilor direct la periost. La sfârșitul fiecărei fibre musculare, plasmolemma formează profunde înguste. Fibrele de colagen subțire le pătrund din tendon sau periost. Acestea din urmă sunt împletite în spirală cu fibre reticulare. Capetele fibrelor sunt îndreptate către membrana subsolului, intră în ea, se întoarce înapoi și, la ieșire, se adună din nou fibrele de colagen ale țesutului conjunctiv.
Între fibrele musculare sunt straturi subțiri de țesut conjunctiv fibros liber - endomisium.Fibrele de colagen ale frunzei exterioare a membranei subsolului sunt țesute în ea (vezi Fig. 9.3, c), care ajută la combinarea eforturilor în timp ce reduc miosimplastele. Straturile mai groase de țesut conjunctiv pierde înconjoară mai multe fibre musculare, formând perimysiumși împărțirea mușchiului în mănunchiuri. Mai multe mănunchiuri sunt combinate în grupuri mai mari, separate de straturi de țesut conjunctiv mai groase. Țesutul conjunctiv care înconjoară suprafața mușchiului se numește epimiziem.
Vascularizare.Arterele intră în mușchi și se răspândesc prin straturile țesutului conjunctiv, subțiri treptat. Ramurile de ordinul al cincilea al șaselea formează arteriole în perimisiu. Capilarele sunt localizate în endomiziu. Ei merg de-a lungul fibrelor musculare, anastomozându-se între ele. Venele, venele și vasele limfatice trec în apropierea navelor de naștere. Ca de obicei, există numeroase mastocite în apropierea vaselor implicate în reglarea permeabilității peretelui vascular.
Inervare.Musculă a evidențiat eferent mielinizat (motor), aferent (sensibil), precum și fibre nervoase vegetative nemielinate. proces celulă nervoasăaducând impuls din neuronul motor măduva spinăriiramuri în perimisia. Fiecare dintre ramurile sale pătrunde prin membrana subsolului și formează terminale la suprafața simplastului de pe plasmolemă, participând la organizarea așa-numitei plăci motorii (vezi cap. 10, Fig. 10.18). La admitere
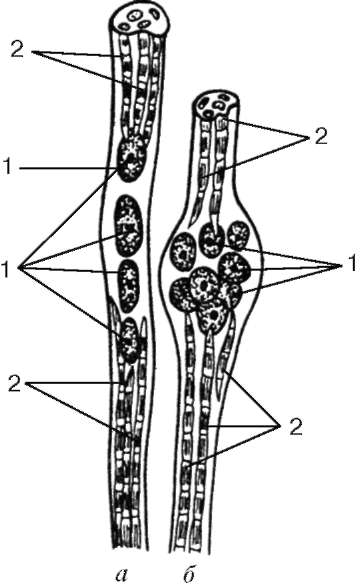
Fig. 9.8.Un fragment dintr-un fus muscular care conține fibre musculare cu un lanț nuclear (a) și cu o pungă nucleară (b) (schemă conform G. S. Katinas): 1 - nuclee; 2 - miofibrilele (nu sunt prezentate organele cu semnificație generală)
acetilcolina este eliberată de la terminalele impulsului nervos - un mediator care provoacă excitație (potențial de acțiune), care se propagă de aici prin plasmolemma miosimplastului.
Deci, fiecare fibră musculară este inervată independent și este înconjurată de o rețea de hemocapilare, formând un complex numit cASE.
Grupul de fibre musculare inervat de un neuron motor este denumit unitate neuromusculară.Fibrele musculare aparținând unei unități neuromusculare nu se află în apropiere, ci sunt situate mozaic printre fibrele aparținând altor unități.
Terminațiile nervoase sensibile nu sunt localizate pe fibrele musculare care funcționează (extrafusal), ci sunt conectate cu fibre musculare specializate în așa-numitele mușchi.
tenakh (cu fibre musculare intrafuzale), care sunt localizate în perimisiu.
Fibrele musculare intrafuzale.Fibrele musculare intrafuzale ale fusurilor sunt mult mai subțiri decât lucrătorii. Există două tipuri: fibra cu pungă nucleară și fibra cu lanț nuclear (Fig. 9.8). Nucleele din cele și din altele sunt rotunjite și localizate în grosimea simplastului și nu la suprafața acestuia. În fibrele cu pungă nucleară, nucleul simplastului formează aglomerații în partea sa mediană îngroșată. În fibrele cu un lanț nuclear în partea de mijloc a simplastului, nu se formează o îngroșare, nucleele se află aici una după alta. Organele de importanță generală sunt situate în apropierea grupurilor de nuclei.
Miofibrilele sunt situate la capetele simplastelor. Fibra de sarcolemă se conectează la capsula fusului neuromuscular, formată dintr-un țesut conjunctiv fibros dens. Fiecare fibră musculară a fusului este legată în spirală cu un terminal al fibrei nervoase senzoriale. Ca urmare a reducerii sau relaxării fibrelor musculare de lucru, tensiunea capsulei de țesut conjunctiv a fusului se modifică, respectiv, tonul fibrelor musculare intrafuziale se modifică. Drept urmare, terminațiile nervoase sensibile sunt excitate în jurul lor și apar impulsuri nervoase aferente în zona terminală. Fiecare miosimplast are, de asemenea, o placă motorie proprie. De aceea, fibrele musculare intrafuzale sunt în continuă tensiune, adaptându-se la lungimea abdomenului muscular în ansamblu.
9.2.3. Țesut muscular muscular
Histogeneză și tipuri de celule.Surse de dezvoltare a țesutului muscular striat cardiac (textus muscularis striatus cardiacus)- secțiuni simetrice ale splanchnotomului frunzei viscerale în partea cervicală a embrionului - plăci mioepicardice.Dintre acestea, mezoteliocitele epicardului se diferențiază și ele. Celulele originale ale țesutului muscular al inimii - explozii cardiomyo- caracterizat printr-o serie de semne: celulele sunt aplatizate, conțin un nucleu mare, un citoplasm ușor, sărac în ribozomi și mitocondrii. În viitor, se dezvoltă complexul Golgi, o rețea endo-plasmatică granulară. Structurile fibrilare se găsesc în cardiomioblaste, dar nu există miofibrilele. Celulele au un potențial proliferativ ridicat.
După o serie de cicluri mitotice, cardiomioblastele se diferențiază în cardiomiocite,în care începe sarcomerogeneza (Fig. 9.9). În citoplasma cardiomiocitelor, numărul polisomilor și tuburilor din reticulul endoplasmic granular crește, se acumulează granule de glicogen și volumul complexului de actomiozină crește. Cardiomiocitele se contractă, dar nu își pierd capacitatea de a prolifera și de a se diferenția în continuare. Dezvoltarea aparatului contractil în perioadele târzii embrionare și postnatală are loc prin adăugarea de noi sarcomere și prin stratificarea miofilamentelor nou sintetizate.
Diferențierea cardiomiocitelor este însoțită de o creștere a numărului de mitocondrii, distribuția lor la poli ai nucleelor \u200b\u200bși între miofibrilele și se desfășoară în paralel cu specializarea suprafețelor de contact ale celulelor. Cardiomiocitele formează contacte „de la capăt”, „de la capăt la altul” pentru a forma fibre musculare cardiace și, în general, țesutul este o structură asemănătoare rețelei. Unele cardiomiocite în primele etape ale cardiomiogenezei sunt contractile-secretorii. Ulterior, ca urmare a diferențierii divergente, apar miocite „întunecate” (contractile) și „ușoare” (conductoare), în care granulele secretoare dispar, în timp ce rămân în miocitele atriale. Deci se formează diferențialul cardiomiocitelor endocrine. Aceste celule conțin un nucleu localizat central cu cromatină dispersată, unul sau doi nucleoli. În citoplasmă este bine dezvoltat un reticul endoplasmic granular, dictiozomi ai complexului Golgi, în strânsă legătură cu elementele cărora se află numeroase granule secretoare cu un diametru de aproximativ 2 μm, care conțin material dens de electroni. Ulterior, granulele secretoare se găsesc sub sarcolemă și sunt secretate în spațiul intercelular prin exocitoză.
În general, cinci tipuri de cardiomiocite apar în timpul histogenezei - de lucru (contractil), sinus (stimulator cardiac), de tranziție, conductor și secretor. Cardiomiocite de lucru (contractile)formează lanțurile lor (fig. 9.10). Atunci când sunt scurtate, acestea asigură forța de contracție a întregului mușchi cardiac. Cardiomiocitele de lucru sunt capabile să
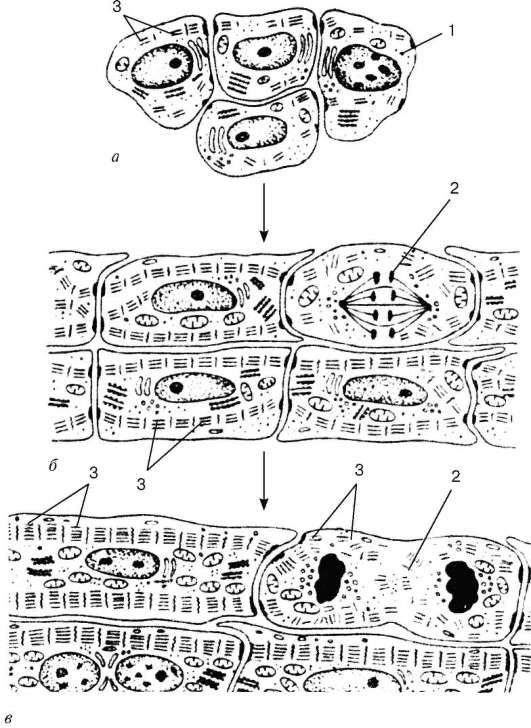
Fig. 9.9.Histogeneza țesutului muscular cardiac (schema conform P.P. Rumyantsev): și- cardiomiocite în peretele tubului cardiac; b -cardiomiocite în embriogeneza târzie; în- cardiomiocite în perioada postanatală. 1 - cardiomiocit; 2 - cardiomiocit care divizează mitotic; 3 - miofilamente și miofibrilele
transmite semnale de control reciproc. Cardiomiocite sinusale (stimulator cardiac)capabil să schimbe automat un anumit ritm starea de contracție la o stare de relaxare. Celulele percep semnale de control din fibrele nervoase, ca răspuns la care schimbă ritmul activității contractile. Cardiomiocitele sinusului (stimulator cardiac) transmit semnale de control cardiomiocite tranzitorii,iar acesta din urmă la cardiomiocite conductoare și de lucru. Conducerea cardiomiocitelorformează lanțuri de celule conectate de capetele lor și sunt situate sub endo-
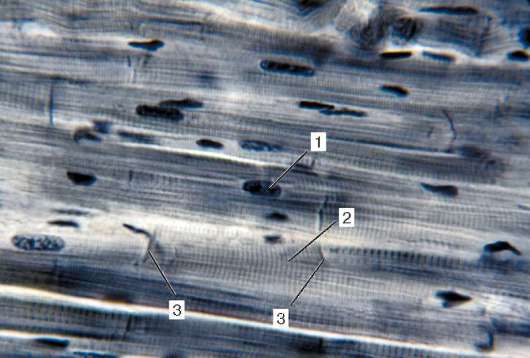
Fig. 9.10.Structura țesutului muscular cardiac (micrografie). Colorare - hematoxilină de fier:
1 - nucleu cardiomiocit; 2 - un lanț de cardiomiocite; 3 - introduceți roți
cardo. Prima celulă din lanț primește semnale de control de la cardiomiocitele sinusale și le transmite altor cardiomiocite conducătoare. Celulele care închid lanțul transmit un semnal prin cardiomiocitele de tranziție către lucrători. Cardiomiocite secretoriiîndeplinește o funcție specială. Produce hormonul peptidic cardiodilatină, care circulă în sânge sub formă de cardionatrina, determină o reducere a arteriolelor miocite netede, o creștere a fluxului sanguin renal și accelerează filtrarea glomerulară și excreția de sodiu. Toate cardiomiocitele sunt acoperite cu o membrană a subsolului.
Structura cardiomiocitelor contractile (de lucru).Celulele au o formă alungită (100-150 μm), aproape de cilindric. Capetele lor sunt conectate între ele, astfel încât lanțurile celulare alcătuiesc așa-numitele fibre funcționale (grosime de până la 20 de microni). În zona contactelor celulare, se formează așa-numitele discuri de inserție (Fig. 9.10). Cardiomiocitele se pot ramifica și forma o rețea spațială. Suprafețele lor sunt acoperite cu o membrană bazală, în care fibrele reticulare și colagen sunt țesute din exterior. Nucleul unui cardiomiocit (uneori două dintre ele) este oval și se află în partea centrală a celulei. La poli din nucleu, sunt concentrate câteva organule de importanță generală, cu excepția reticulului endoplasmatic și a mitocondriilor agranulare.
Organele speciale care asigură contracția se numesc miofibrilele. Sunt slab izolate unele de altele, pot fi împărțite. Structura lor este similară cu structura miofibrilelor miosimplastului fibrei musculare scheletice. Fiecare mitocondrie este localizată în întregul sarcomere. Tuburile T situate la nivelul liniei Z sunt direcționate de la suprafața membranei plasmatice în adâncimea cardiomiocitului. Membranele lor sunt reunite
în contact cu membranele rețelei endoplasmice (sarcoplasmatice) netede. Buclele din acestea din urmă sunt alungite de-a lungul suprafeței miofibrilelor și au extensii laterale (sisteme L), care formează, împreună cu tuburile T, o triadă sau o diadă (Fig. 9.11, a). În citoplasmă sunt incluse glicogenul și lipidele, în special multe incluziuni ale mioglobinei. Mecanismul de contracție a cardiomiocitelor este același ca în miosimplast.
Organizarea cardiomiocitelor în țesut.Cardiomiocitele sunt conectate între ele prin tipul de „capăt la capăt”. Discurile de inserție se formează aici: aceste zone arată ca niște plăci subțiri cu o mărire medie a microscopului ușor. De fapt, capetele cardiomiocitelor au o suprafață neuniformă, astfel încât proeminențele unei celule intră în jgheaburile alteia. Secțiunile transversale ale proeminențelor celulelor adiacente sunt interconectate prin interdigitații și desmosomi (Fig. 9.11, b).
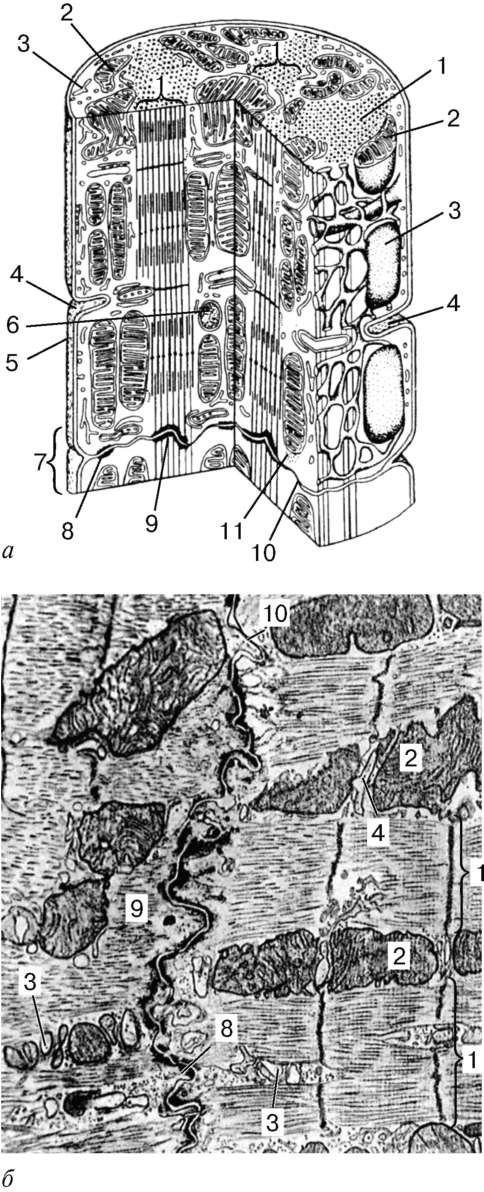
Fig. 9.11.Structura cardiomiocitului: și- schema (conform Yu. I. Afanasyev și V. L. Goryachkina); b- micrografia electronică a discului de inserție. O creștere de 20 000. 1 - miofibrilele; 2 - mitocondrii; 3 - rețea sarcotubulară; 4 - tuburi T; 5 - membrană subsol; 6 - lizozom; 7 - introduceți disc; 8 - desmosome; 9 - zona de atașare a miofibrilelor; 10 - contacte cu sloturi; 11 - glicogen
La fiecare desmosome din partea citoplasmei, miofibril la se apropie, care este fixat până la capăt în complexul desmo-placă. Astfel, în timp ce reducerea poftei unui cardiomiocit este transmisă altuia. Suprafețele laterale ale proeminențelor cardiomiocitelor sunt unite de nexuzi (joncțiuni la distanță). Aceasta creează legături metabolice între ele și asigură sincronizarea contracțiilor.
Regenerarea.În histogeneza țesutului muscular cardiac, cambiumul nu apare. Prin urmare, regenerarea țesuturilor are loc pe baza proceselor hiperplastice intracelulare. În același timp, procesul de poliploidie este caracteristic pentru cardiomiocitele mamiferelor, primatelor și oamenilor.
tION. De exemplu, la maimuțe, până la 50% din cardiomiocitele diferențiate în nucleu devin tetra- și octoploide. Cardiomiocitele poliploide apar din cauza mitozei acitokinetice, ceea ce duce la multinucleare. În condițiile patologiei sistemului cardiovascular uman (reumatism, defecte cardiace congenitale, infarct miocardic etc.), regenerarea intracelulară, poliploidizarea nucleelor \u200b\u200bși apariția cardiomiocitelor multinucleate joacă un rol important în compensarea afectării cardiomiocitelor.
9.3. FABRICI MUSCULARE SCURT
Există trei grupuri de țesut muscular neted (fără striat) (textus muscularis nonstriatus)și celule: celenele mezenchimale, neuronale și mioepiteliale.
9.3.1. Țesut muscular de origine mezenchimală
Histogeneză.Acest țesut este împărțit în două tipuri: viscerale și vasculare. În histogeneza embrionară, chiar și microscopia electronică este greu de diferențiat între progenitorii fibroblastului mezenchimal și miocitele netede. În miocitele netede slab diferențiate, se dezvoltă un reticul endoplasmic granular, complexul Golgi. Filamentele subțiri sunt orientate de-a lungul axei lungi a celulei. Pe măsură ce se dezvoltă, mărimile celulelor și numărul de filamente din citoplasmă cresc. Treptat, volumul citoplasmei ocupate de filamentele contractile crește, localizarea lor în citoplasmă devine tot mai ordonată. Activitatea proliferativă a miocitelor netede în miogeneză este redusă treptat. Aceasta apare ca urmare a creșterii duratei ciclului celular, a ieșirii celulelor din ciclul reproducător și a tranziției la o stare diferențiată. Diferențindu-se, acestea sintetizează componentele matricei intercelulare, colagenul membranei subsolului, precum și elastina. În celulele definitive (miocite), capacitatea sintetică este redusă, dar nu dispare complet.
Structura și funcționarea celulelor.Un miocit neted este o celulă în formă de fus lung de 20-500 microni, lățime de 5-8 microni. Miezul are formă de tijă, situat în partea sa centrală. Când miocitul se contractă, miezul său se îndoaie și chiar se răsucește (Fig. 9.12–9.14).
Structura miocitelor netede definitive (leiomiocite) care alcătuiesc organele interne și pereții vaselor de sânge are multe în comun, dar în același timp este caracterizată de heteromorfism. Așadar, în pereții venelor și arterelor se găsesc miocite procesoare în formă de fus, cu o lungime de 10-40 microni, ajungând uneori până la 140 microni. Cea mai lungă miO
cytes ajung până la 500 de microni în peretele uterin. Diametrul miocitelor este cuprins între 2 și 20 microni. În funcție de natura proceselor biosintetice intracelulare, se disting miocite contractile și secretorii. Primele sunt specializate în funcția de contracție, dar, în același timp, păstrează activitatea secretorie.
Miocitele secretoare din ultrastructura lor seamănă cu fibroblaste, dar conțin în citoplasmă mănunchiuri de miofilamente subțiri situate la periferia celulei. Complexul Golgi, reticulul endoplasmatic granular, multe mitocondrii, granule glicogene, ribozomi liberi și polisomi sunt bine dezvoltați în citoplasmă. După gradul de maturitate, astfel de celule sunt clasificate ca fiind slab diferențiate. Filamentele de actină din citoplasmă formează o rețea tridimensională, alungită în principal longitudinal. Capetele filamentelor sunt legate între ele și plasmolemmei cu proteine \u200b\u200breticulante speciale. Aceste zone sunt clar vizibile în micrografele electronice sub formă de corpuri dense. Monomerii de miozină sunt localizați lângă filamentele de actină. Plasmolemma formează intruziuni - caveole, în care ionii de calciu sunt concentrați. Semnalul contracției vine de obicei prin fibrele nervoase. Mediatorul, care se evidențiază din termenii lor, schimbă permeabilitatea plasmolemmei. Ionii de calciu sunt eliberați, ceea ce implică atât polimerizarea miozinei, cât și interacțiunea miozinei cu actina.
Există o retragere de actină-miofilamente între mioză
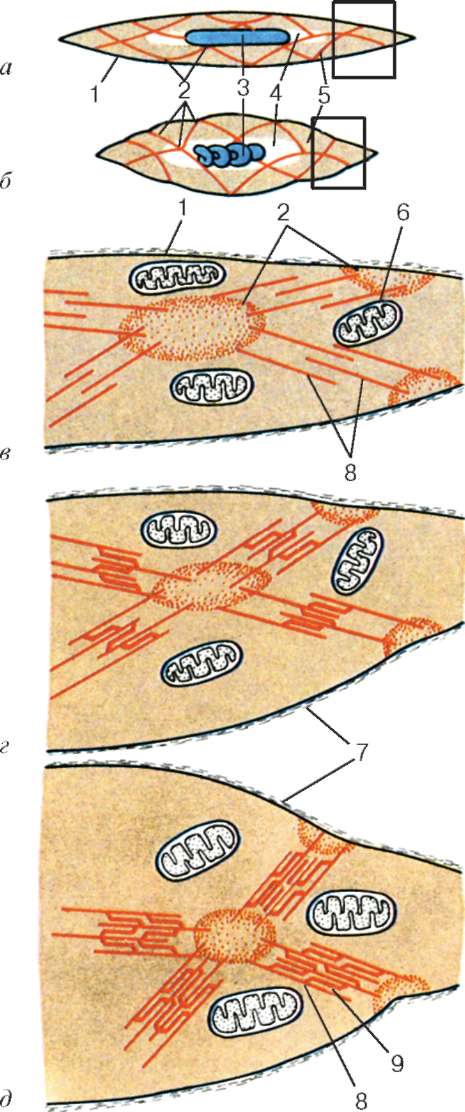
Fig. 9.12.Structura unui miocit neted (schemă):
și, în- la relaxare; b. d- la cea mai mare reducere; g- cu reducere incompletă; in-d- imagini mărite ale secțiunilor încadrate pe fragmente șiși b. 1 - plasmolemă; 2 - corpuri dense; 3 - miez; 4 - endoplasmă; 5 - complexe contractile; 6 - mitocondrii; 7 - membrana subsolului; 8 - miofilamente de actină (subțiri); 9 - miofilamente (groase) de miozină
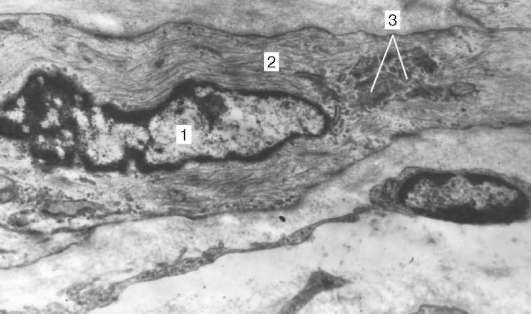
Fig. 9.13.Ultrastructură de diferențiere a miocitului neted în peretele bronhiei:
1 - miez; 2 - citoplasmă cu miofilamente; 3 - Complexul Golgi, o creștere de 35 000 (medicament A. L. Zashikhina)
se întâlnesc pete noi și dense, forța este transmisă plasmolemmei și întreaga celulă este scurtată (vezi Fig. 9.12). Când semnalele sistemului nervos încetează, ionii de calciu se deplasează de la citoplasmă la caveola și tubulele reticulului endoplasmic, miozina este depolimerizată și miofibrilele se rup. Contracția se oprește. Astfel, complexele de actinomiozină există în miocite netede numai în perioada de contracție, în prezența ionilor de calciu liberi în citoplasmă.
Miocitele sunt înconjurate de o membrană a subsolului. În anumite zone, în el se formează „ferestre”, astfel încât plasmolemma miocitelor vecine converg. Aici, se formează nexuzele și nu numai mecanice, ci și legături metabolice apar între celule. În partea de sus a „copertelor” de la membrana subsolului dintre miocite sunt fibre elastice și reticulare, care combină celulele într-un singur complex tisular. Miocitele netede sintetizează proteoglicani, glicoproteine, procolagen, pro-elastină, din care se formează fibre de colagen și elastice și componenta amorfă a matricei intercelulare. Interacțiunea miocitelor se realizează cu punți citoplasmatice, proeminențe reciproce, nexuze, desmosomi, secțiuni de contacte ale membranelor suprafețelor miocitelor.
Regenerarea.Țesutul muscular neted al speciilor viscerale și vasculare are sensibilitate semnificativă la efectele factorilor extreme. În miocitele activate, nivelul proceselor biosintetice crește, a cărui expresie morfologică este sinteza proteinelor contractile, extinderea și hipercromatoza nucleului, hipertrofia nucleului, o creștere a raportului citoplasmatic nuclear, o creștere a numărului de ribozomi și polisomi liberi și

Fig. 9.14.Structura țesutului muscular neted (schema volumetrică) (conform R.V. Krstic, cu modificări):
1 - miocite netede fusiforme; 2 - citoplasma miocitului; 3 - nuclee de miocite; 4 - plasmolemă; 5 - membrană subsol; 6 - vezicule pinocitotice superficiale; 7 - conexiuni intercelulare; 8 - terminarea nervului; 9 - fibrilele de colagen; 10 - microfilamente
enzimele, fosforilarea aerobă și anaerobă, transportul membranei. Regenerarea celulară se realizează atât datorită celulelor diferențiate cu capacitatea de a intra în ciclul mitotic, cât și datorită activării elementelor cambiale (miocite de volum mic). Sub acțiunea mai multor factori dăunători, se observă o transformare fenotipică a miocitelor contractile în secretoare. Această transformare este adesea observată cu deteriorarea intimei vasculare, formarea hiperplaziei intime cu dezvoltarea aterosclerozei.
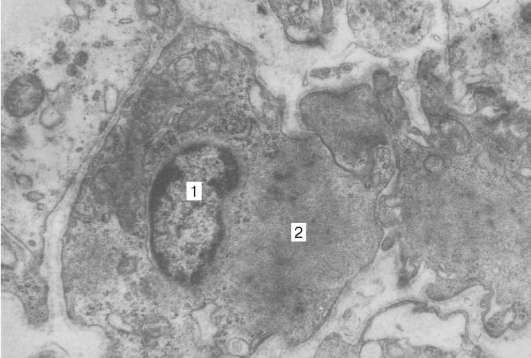
Fig. 9.15.Ultrastructura miopigmentocitului (medicament N. N. Sarbaeva): 1 - nucleu; 2 - miofilamente, o creștere de 6000
9.3.2. Țesutul muscular mezenchimal în organe
Miocitele sunt combinate în mănunchiuri, între care există straturi subțiri de țesut conjunctiv. Fibrele reticulare și elastice din jurul miocitelor sunt țesute în aceste straturi. Vasele de sânge și fibrele nervoase trec prin straturile intermediare. Terminalele acestora din urmă nu se termină direct pe miocite, ci între ele. Prin urmare, după sosirea unui impuls nervos, mediatorul se răspândește difuz, excitând multe celule simultan. Țesutul muscular neted de origine mezenchimală este reprezentat mai ales în pereții vaselor de sânge și în multe organe interne scobite.
Țesutul muscular neted în anumite organe are proprietăți funcționale inegale. Acest lucru se datorează faptului că pe suprafața organelor există receptori diferiți pentru substanțe biologice specifice biologic. Prin urmare, pentru multe medicamente, reacția lor nu este aceeași. Este posibil ca diferite proprietăți funcționale ale țesuturilor să fie asociate cu o organizare moleculară specifică a filamentelor de actină.
9.3.3. Țesut muscular de origine neurală
Țesutul muscular al irisului și corpului ciliar este al patrulea tip de țesut contractil. Miocitele acestui țesut se dezvoltă din celulele rudimentului neural ca parte a peretelui intern al gobletului optic. La rând
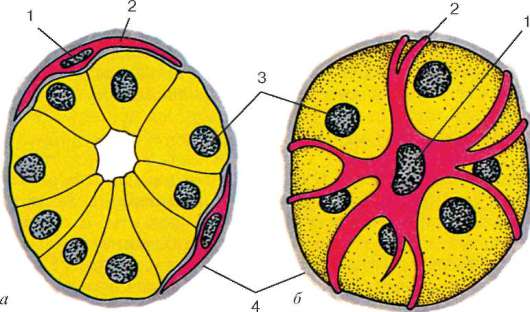
Fig. 9.16.Celulele mioepiteliale în secțiunea finală a glandei salivare (schema conform G. S. Katinas):
și- secțiune transversală; b- vedere de la suprafață. 1 - nucleii celulelor mioepiteliale; 2 - procese ale celulelor mioepiteliale; 3 - nuclee de celule epiteliale secretoare; 4 - membrană subsol
elementele musculare vertebrale ale irisului prezintă o diferențiere divergentă. Astfel, țesutul mioneural la reptile și păsări este reprezentat de fibre multinucleate striate, care sunt foarte asemănătoare cu mușchiul scheletului. La mamifere și oameni, principala unitate structurală și funcțională a mușchilor irisului este un miocit mononuclear neted sau miopigmentocit. Acestea din urmă au un corp pigmentat care conține un miez, scos în afara părții contractile în formă de fus (Fig. 9.15).
Citoplasma celulelor conține un număr mare de mitocondrii și granule de pigment, care sunt similare ca mărime și formă cu granulele epiteliului pigmentar. Miofilamentele din miopigmentocite sunt împărțite în subțiri (7 nm) și groase (1,5 nm), ca mărime și locație, seamănă cu miofilamente de miocite netede. Fiecare miopigmentocit este înconjurat de o membrană a subsolului. În apropierea proceselor citoplasmatice ale miocitelor, se găsesc fibre nervoase fără mielină. În funcție de direcția proceselor (perpendicular sau paralel cu marginea pupilei), miocitele formează doi mușchi - o pupilă care se îngustează și se extinde.
Regenerarea.Câteva studii au arătat o activitate redusă de regenerare după deteriorare sau absența acesteia.
9.3.4. Celulele musculare epidermice
Celulele mioepiteliale se dezvoltă din germenul epidermic. Se găsesc în glande sudoripare, mamare, salivare și lacrimale și au precursori comuni cu celulele lor secretoare. mioepiteliale
celulele sunt direct adiacente epiteliului propriu-zis și au o membrană comună a subsolului cu ele. În timpul regenerării, celulele și alte celule sunt, de asemenea, restaurate de la precursorii comuni cu diferențe reduse. Majoritatea celulelor mioepiteliale au formă de stea. Aceste celule sunt adesea numite în formă de coș: procesele lor acoperă secțiunile de capăt și conductele mici ale glandelor (Fig. 9.16). Nucleul și organulele de importanță generală sunt localizate în corpul celulei, iar aparatele contractile, organizate ca în celulele țesutului muscular de tip mezenchimal, sunt localizate în procese.
Întrebări de securitate
1. Clasificarea genetică a țesutului muscular. Unități structurale și funcționale ale diferitelor tipuri de țesut muscular.
2. Țesut muscular muscular scheletat: dezvoltare, structură, bază morfologică a contracției musculare. Regenerarea.
3. Țesut muscular muscular striat: dezvoltare, structură specifică a diferitelor tipuri de cardiomiocite, regenerare.
4. Soiuri de miocite netede: surse de dezvoltare, topografie în organism, regenerare.
Histologie, embriologie, citologie: manual / Yu. I. Afanasyev, N. A. Yurina, E. F. Kotovsky ș.a. - ediția a 6-a, Rev. și adăugați. - 2012 .-- 800 s. : bolnav.
Mușchii corpului uman sunt formați în principal din țesut muscular, format din celule musculare. Distingeți între țesutul muscular neted și striat. Țesut muscular fin ea construiește musculatura, care face parte din unele organe interne și striatformează mușchiul scheletului. O proprietate comună a țesutului muscular este excitabilitate, conductibilitate și contractilitatea.
Țesutul muscular striat se distinge de excitabilitate, conductivitate și contractilitate netedă. Celulele mușchiului striat au un diametru foarte mic și o lungime mare (până la 10-12 cm). În acest sens, sunt numiți fibre.
La fel ca alte celule, celulele musculare au un protoplasm numit sarcoplasma(din greacă. Sarcos - carne). Numită membrană celulară musculară sarcolemă. În interiorul fibrei musculare sunt numeroși nuclei și alte componente ale celulelor.
Compoziția fibrelor musculare include un număr mare de fibre și mai subțiri - myofibrils, care, la rândul lor, constau din cele mai subțiri fire - protofibrilele. protofibril - acesta este aparatul contractil al celulei musculare, sunt proteine \u200b\u200bcontractile speciale - miozina și actina. Mecanismul contracțiilor musculare este un proces complex de transformări fizice și chimice în fibra musculară, cu participarea obligatorie a aparatului contractil. Acest mecanism este declanșat de un impuls nervos, iar energia pentru procesul de reducere este furnizată de acidul adenosin trifosforic (ATP). În acest sens, o caracteristică a structurii fibrelor musculare este, de asemenea, un număr mare de mitocondrii care asigură fibrei musculare energia necesară. Relaxarea fibrelor musculare, după mulți oameni de știință, se realizează pasiv, datorită elasticității sarcolemmei și a țesutului conjunctiv intramuscular.
Structura, forma și clasificarea mușchiului scheletului
Unitatea anatomică a celei mai active părți a sistemului muscular muscular - mușchii scheletici sau striați - este mușchiul scheletului. Mușchiul scheletului - Acesta este un organ format din țesut muscular muscular striat, care conține și țesut conjunctiv, nervi și vase de sânge.
Fiecare mușchi este înconjurat de un fel de „caz” de țesut conjunctiv - fasciei.
în structură externă mușchii disting între capul tendonului corespunzător începutului mușchiului, abdomenului mușchiului sau corpul format din fibre musculare și capătul tendonului mușchiului sau coada, cu care mușchiul se atașează de un alt os. De obicei, coada mușchiului este un punct de mișcare de atașare, iar începutul este fixat. În procesul mișcării, funcțiile lor se pot schimba: punctele de mișcare devin nemișcate și invers.
Forma mușchilor este foarte diversă și depinde în mare măsură de scopul funcțional al mușchiului. Distingeți între mușchii lungi, scurti, largi, circulari și alți mușchi. lung mușchii sunt localizați pe membre scurt - acolo, unde mișcarea este mică (de exemplu, între vertebre). Mușchii largi sunt localizați în principal pe corp (mușchii abdomenului, spatelui, pieptului). Mușchii circulari sunt localizați în jurul deschiderilor corpului și îi restricționează atunci când se contractă. Acești mușchi sunt numiți sfinctere. Dacă un mușchi are un cap, se numește simplu, dacă sunt două sau mai multe - complexe (de exemplu, biceps, triceps și cvadriceps). O trăsătură morfologică importantă este localizarea fibrelor musculare. Există aranjamente paralele, oblice, transversale și circulare ale fibrelor (în sfinctere). dacă, cu un aranjament oblic al fibrelor musculare, acestea se atașează doar pe o parte la tendoane, atunci mușchii sunt numiți semipenniformdacă de ambele părți - dvuperistymi.
Funcțional, mușchii pot fi împărțiți în flexorii și extensor, rotatoare exterioare și rotitoare în interior, mușchi adductor și vents. Muschii sunt secretati si - sinergici și mușchii - antagoniști. Primii formează un grup de mușchi, care efectuează cu ușurință orice mișcare, contracția acestuia din urmă provoacă mișcări opuse.
Grupuri musculare majore
Mușchii scheletici ai unui adult alcătuiesc aproximativ 40% din greutatea corpului său. La nou-născuți și copii mici, mușchii nu reprezintă mai mult de 20-25% din greutatea corporală, iar la bătrânețe există o scădere treptată a masei musculare până la 25-30% din greutatea corporală. În total, corpul uman are peste 600 de mușchi.
După locație, adică pe motive topografice se disting mușchii spatelui, toracelui, abdomenului, capului, gâtului, extremităților superioare și inferioare.
Mușchii spatelui sunt împărțiți în suprafață și adâncime. K mușchii superficiali spatele include: mușchiul trapez, latissimus dorsi, mușchii romboizi (major și minor), mușchii dentari (superior și inferior).
Acești mușchi sunt implicați în mișcările membrelor superioare și în modificările de volum. torace (act de respirație).
Mușchii adânci spatele este reprezentat de numeroși mușchi localizați de-a lungul coloanei vertebrale. Sunt atașate în principal de procesele vertebrelor și sunt implicați în mișcările coloanei vertebrale înapoi și în lateral.
Mușchii pieptului sunt împărțiți în mușchi aparținând brâului de umăr și membrul superior (mușchii pectorali majori și minori, subclavieni și mușchiul dentat anterior) , și mușchii proprii ai toracelui (mușchii intercostali interni externi). Se pun în mișcare și întăresc membrele superioare ale corpului și participă la funcția de respirație. Diafragma aparține și mușchilor respiratorii (separă cavitatea toracică de cavitatea abdominală).
Mușchii abdominali sunt reprezentați de mușchii abdominis oblici, transversi și recti, externi și interni, precum și de mușchiul pătrat din partea inferioară a spatelui.
Mușchii rectului abdominis participă la îndoirea înainte a corpului, iar mușchii oblici oferă înclinarea coloanei vertebrale către părțile laterale și rotirile acesteia împreună cu pieptul la dreapta și la stânga. Mușchii abdominali nu participă numai la mișcările trunchiului și pieptului. Ele formează un zid cavitatea abdominală și, datorită tonului lor, își păstrează organele interne în poziția normală. Prin reducerea acestora, cresc presiunea intraabdominală, formând așa-numita abs. Mușchii abdominali facilitează mișcările intestinale (mișcări ale intestinului), urinarea, iar la femei expulzarea fătului în timpul nașterii. Contracția mușchilor abdominali contribuie, de asemenea, la mișcarea sângelui în sistemul venos și la implementarea mișcărilor respiratorii, dar și la formarea unei hernii. Cu o hernie, apare o proeminență a organelor interne (intestine, stomac etc.) din cavitatea abdominală sub piele.
Printre mușchii capului, se disting mușchii mestecători și mușchii faciali. K mesteca muschii se aplică temporal, mestecat, pterygoid. Contracția acestor mușchi determină mișcări complexe de mestecare. maxilarul inferior. Mușchii mimici diferă de alți mușchi ai corpului uman prin faptul că sunt atașați de pielea feței cu unul, și uneori cu două, de capetele lor. Contracția acestor mușchi este deplasarea pielii, formarea pliurilor și a ridurilor. Aceasta definește expresiile faciale, adică una sau alta expresie facială. Manifestarea unor senzații complexe (emoții) - bucurie, rușine, dispreț, durere și durere etc. - Este determinat de numeroase combinații de contracții ale mușchilor faciali, subordonate impulsurilor care le duc din cortexul cerebral de-a lungul nervului facial. Mușchii circulari ai ochiului și ai gurii sunt, de asemenea, imitați. Mușchii gâtului înclină capul, îl înclină și se întoarce. Mușchii atașați de osul hioid, în timpul contracției, schimbă poziția limbii și a laringelui atunci când înghiți și scoate sunete.
Mușchii membrului superior sunt împărțiți în mușchi brâu de umăr și mușchii membrului superior liber. Mușchii brâului umărului înconjoară articulația umărului, oferind numeroase mișcări în ea. Printre ei, cel mai important este mușchiul deltoid. Când se contractă, acest mușchi îndoaie brațul articulația umărului și își duce mâna în poziția orizontală.
Mușchii membrului superior liber includ mușchii umărului, antebrațului și ai mâinii.
În zona umărului din față este o grupare musculară - flexoriiînapoi - extensor. Dintre mușchii grupului anterior, cel mai important este mușchiul biceps al umărului (biceps), iar spatele, mușchiul triceps al umărului.
Mușchii antebrațului sunt reprezentați și pe suprafața frontală de către flexori, iar pe spate de extensori. Mușchii mâinii sunt localizați doar pe suprafața palmarului. Printre ele ies în evidență - un mușchi palmar lung și flexorii degetelor. Datorită prezenței propriului sistem muscular, în special, degetele deget mare, dobândesc o mobilitate mai mare și sunt capabili de diverse mișcări, inclusiv flexie și extensie maximă, ceea ce este extrem de important atunci când lucrați.
Mușchii situați în regiunea centurii extremității inferioare mută piciorul înăuntru articulația șoldului, precum și cu un membru fix, acestea îndoaie coloana vertebrală în regiunea lombară. Grupul muscular anterior include un mușchi major - iliopsoas. Printre grupul extern posterior de mușchi al brâului pelvin - mușchii gluteali mari, mijlocii și mici.
Membre inferioare au un schelet mai masiv decât cele superioare; mușchii lor au o mare rezistență, dar, în același timp, o varietate mai mică și o gamă limitată de mișcare.
Pe coapsa din față este cel mai lung mușchi croitor din corpul uman (până la 50 cm). Îndoaie piciorul în articulațiile șoldului și genunchiului.
Mușchiul quadriceps femoris se află mai adânc decât mușchiul croitor, încercuind femurul din aproape toate părțile.
Principala funcție a acestui mușchi este extinderea articulației genunchiului. Când stă în picioare, cvadricepsul nu dă articulația genunchiului a se apleca.
Pe picior se disting mușchii spatelui și plantar. Acești mușchi sunt implicați în flexia și extinderea degetelor de la picioare, precum și în convergența și reproducerea lor.