Describa los principales tipos de tejido muscular. La estructura histológica del tejido muscular.
El tejido muscular se llama tejido que tiene una estructura y un origen diferentes, pero una capacidad similar para contracciones pronunciadas. Proporcionan movimiento en el espacio de todo el organismo como un todo o sus partes (por ejemplo, músculos esqueléticos) y el movimiento de los órganos dentro del órgano (por ejemplo, el corazón, la lengua, los intestinos).
Las células de muchos tejidos poseen la propiedad del cambio de forma, pero esta capacidad se convierte en la función principal del tejido muscular.
Característica general y clasificación
Los principales signos morfológicos de los elementos del tejido muscular son la forma alargada, la presencia de miofibrillas y miofilamentos ubicados longitudinalmente, orgánulos especiales que proporcionan contractilidad, la ubicación de las mitocondrias cerca de elementos contráctiles, la presencia de inclusiones de glucógeno, lípidos y mioglobina.
Tejido muscular liso.
Este tejido está formado por mesénquima. Células del músculo liso: miocitos alargados con extremos puntiagudos fusiformes, cubiertos con una membrana celular. Un miocito es una célula mononuclear. El núcleo en forma de barra ocupa una posición central en la célula. Contiene heterocromatina atomizada y uno o dos nucleolos claramente visibles. El centro celular (citocentro) se encuentra cerca de una de las superficies del núcleo. Los organelos restantes se concentran en los polos del núcleo en forma de barra. Mitocondrias bien desarrolladas, complejo de Golgi, EPS liso; ribosomas, EPS granular se expresan ligeramente. El citoplasma de los miocitos contiene gránulos de glucógeno, la reserva de energía de la célula.
Los miocitos están estrechamente adyacentes entre sí, formando capas y grupos, separados por un tejido conectivo suelto sin forma. Las miofibrillas se encuentran en el citoplasma, van a lo largo de la periferia de la célula a lo largo de su eje. Consisten en hilos delgados y son un elemento contráctil del músculo.
En el exterior, la fibra muscular está cubierta con una membrana, un sarcolema, que consta de las capas interna y externa. La capa interna es un plasmolemma, que es similar a la membrana de otras células de tejido. Exterior: la capa de tejido conjuntivo consiste en una membrana basal y estructuras fibrosas adyacentes. El plasmolema forma un sistema de túbulos estrechos que penetran en la fibra muscular.
Las fibras del tejido conectivo ubicadas fuera de la membrana basal de la fibra muscular forman un endomisio, que es rico en vasos sanguíneos y nervios. El endomisio se combina con el perimisio, una membrana que cubre un grupo de fibras musculares. El perimisio de varios haces musculares está conectado con el epimisio, la membrana más externa del tejido conectivo que combina varios de estos haces en un músculo, un órgano caracterizado por una estructura y función específicas.
El tejido muscular liso se encuentra en las paredes del estómago, intestinos, útero, vejiga, uréteres, bronquios, así como en vasos sanguíneos medianos y grandes. La actividad del músculo liso está regulada por el ANS. Las contracciones musculares no obedecen la voluntad de una persona y, por lo tanto, el tejido muscular liso se llama musculatura involuntaria.
Tejido muscular estriado.
El tejido muscular esquelético es el tejido contráctil del tronco, la cabeza, las extremidades, la faringe, la laringe, la mitad superior del esófago, la lengua y los músculos masticatorios.
Se forma a partir de los miotomas de los somitas del mesodermo. La unidad estructural es la fibra muscular estriada. Tiene un cuerpo cilíndrico, está cubierto con una concha (sarcolemo) y el citoplasma se llama sarcoplasma, en el que hay numerosos núcleos y miofibrillas. Cada miofibril consiste en discos que tienen una composición química diferente. Las miofibrillas son el aparato contráctil de la fibra muscular.
Todos los músculos esqueléticos se construyen a partir del tejido muscular estriado. Los músculos son arbitrarios, porque su contracción ocurre bajo la influencia de la corteza cerebral.
Tejido muscular del corazón.
Este tipo de tejido muscular forma la membrana media del corazón: el miocardio, por la naturaleza de la reducción, se refiere a involuntario, ya que no está controlado por la voluntad del animal. Se desarrolla a partir del esplácnatoma de la hoja visceral, lámina mioepicárdica.
El tejido muscular cardíaco consta de células musculares: cardiomiocitos (miocitos cardíacos). Los miocitos, que se conectan entre sí en sus extremos a lo largo del eje longitudinal de las células, forman una estructura similar a la fibra muscular.
Hay dos tipos de células. : células contráctiles típicas y miocitos cardíacos atípicosconstituyendo el sistema de conducción del corazón.
Células musculares típicas realizar la función contráctil; son de forma rectangular, 1-2 núcleos están en el centro, miofibrillas se encuentran en la periferia. Entre los miocitos adyacentes hay discos de inserción. Con su ayuda, los miocitos se acumulan en fibras musculares, separadas por un tejido conectivo fibroso y delgado. Entre las fibras musculares adyacentes hay fibras de conexión que proporcionan la contracción del miocardio en su conjunto.
El sistema de conducción del corazón está formado por fibras musculares que consisten en células musculares atípicas. Son más grandes que contráctiles, más ricos en sarcoplasma, pero más pobres que las miofibrillas, que a menudo se cruzan. Los granos son más grandes y no siempre están en el centro. Las fibras del sistema conductor están rodeadas por un denso plexo de fibras nerviosas.
Los tejidos musculares (lat. Textus muscularis) se denominan tejidos que tienen una estructura y un origen diferentes, pero una capacidad similar para contracciones pronunciadas. El tejido muscular está formado por células alargadas que toman irritación de sistema nervioso y responde con una reducción. Proporcionan movimiento en el espacio del cuerpo en su conjunto, su movimiento de órganos dentro del cuerpo (corazón, lengua, intestinos, etc.) y consisten en fibras musculares. Las células de muchos tejidos poseen la propiedad del cambio de forma, pero en los tejidos musculares esta capacidad se convierte en la función principal.
Los principales signos morfológicos de los elementos del tejido muscular: una forma alargada, la presencia de miofibrillas y miofilamentos ubicados longitudinalmente, orgánulos especiales que proporcionan contractilidad, la ubicación de las mitocondrias cerca de elementos contráctiles, la presencia de inclusiones de glucógeno, lípidos y mioglobina.
Los orgánulos contráctiles especiales, miofilamentos o miofibrillas, proporcionan la contracción que ocurre cuando dos proteínas fibrilares principales, actina y miosina, interactúan con ellas, con la participación obligatoria de iones de calcio. Las mitocondrias proporcionan energía a estos procesos. El stock de fuentes de energía está formado por glucógeno y lípidos. La mioglobina es una proteína que proporciona la unión del oxígeno y la creación de su reserva en el momento de la contracción muscular, cuando se comprimen los vasos sanguíneos (el suministro de oxígeno cae bruscamente).
Propiedades del tejido muscular:
- Excitabilidad
- Conductividad
- Contractilidad
- Labilidad
Tipos de tejido muscular:
1. Tejido muscular liso
El tejido muscular liso consiste en células mononucleares, miocitos en forma de huso con una longitud de 20-500 micras. Su citoplasma en un microscopio óptico se ve uniforme, sin estriación transversal. Este tejido muscular tiene propiedades especiales: se contrae y relaja lentamente, tiene automatización, es involuntario (es decir, su actividad no está controlada por la voluntad de una persona). Incluido en las paredes órganos internos: vasos sanguíneos y linfáticos, tracto urinario, tracto digestivo (contracción de las paredes del estómago y los intestinos).
2. Tejido estriado del músculo esquelético
Consiste en miocitos que tienen una gran longitud (hasta varios centímetros) y un diámetro de 50-100 micras; estas células son multinucleadas, contienen hasta 100 o más núcleos; En un microscopio óptico, el citoplasma parece una alternancia de rayas oscuras y claras. Las propiedades de este tejido muscular son una alta tasa de contracción, relajación y aleatoriedad (es decir, su actividad está controlada por la voluntad de una persona). Este tejido muscular es parte de los músculos esqueléticos, así como las paredes de la faringe, la parte superior del esófago, forma la lengua, los músculos oculomotores. Fibras de 10 a 12 cm de largo.
3. Tejido muscular cardíaco estriado
Consiste en 1 o 2 cardiomiocitos nucleares que tienen una estriación transversal del citoplasma (a lo largo de la periferia del citolema). Los cardiomiocitos son ramificados y forman conexiones interconectadas: discos de inserción en los que se combina su citoplasma. También hay otro contacto intercelular, las anostamosis (la inhibición del citoplomama de una célula en el citomalma de otra). Este tipo de tejido muscular forma el miocardio del corazón. Se desarrolla a partir de la placa mioepicárdica (hoja visceral del esplnototoma del cuello del embrión). Una propiedad especial de este tejido es la automatización: la capacidad de contraerse y relajarse rítmicamente bajo la influencia de la excitación que ocurre en las células (cardiomiocitos típicos). Este tejido es involuntario (cardiomiocitos atípicos). Existe un tercer tipo de cardiomiocito: los cardiomiocitos secretores (no tienen fibrillas), que sintetizan la hormona troponina, que disminuye la presión arterial y expande las paredes de los vasos sanguíneos.
La membrana muscular del corazón (miocardio) está formada por células de la cavidad muscular, que están conectadas a complejos musculares o fibras musculares cardíacas con la ayuda de discos de inserción. El tejido muscular tiene la capacidad de recuperarse, está protegido por tejido conectivo, formando una cicatriz.
Presentar su buen trabajo a la base de conocimiento es fácil. Use el formulario a continuación
Los estudiantes, los estudiantes de posgrado, los jóvenes científicos que utilizan la base de conocimientos en sus estudios y trabajos le estarán muy agradecidos.
Publicado en http://www.allbest.ru/
Tejido muscular combina la capacidad de reducir.
Características estructurales: el aparato contráctil, que ocupa una parte importante en el citoplasma de los elementos estructurales del tejido muscular y consiste en filamentos de actina y miosina, que forman orgánulos especiales. miofibrillas .
Tejido muscularson un grupo de tejidos de diversos orígenes y estructuras, unidos sobre la base de una característica común: una capacidad contráctil pronunciada, debido a la cual pueden realizar su función principal, para mover el cuerpo o sus partes en el espacio.
Las propiedades más importantes del tejido muscular.Los elementos estructurales del tejido muscular (células, fibras) tienen una forma alargada y son capaces de contraerse debido al poderoso desarrollo del aparato contráctil. Este último se caracteriza por una disposición altamente ordenada actinay miofilamentos de miosina,creando condiciones óptimas para su interacción. Esto se logra uniendo estructuras contráctiles con elementos especiales del citoesqueleto y el plasmolema (sarcolema)realizando función de referencia. En la parte del tejido muscular, los miofilamentos forman orgánulos de especial importancia: miofibrillasPara la contracción muscular, se requiere una cantidad significativa de energía, por lo tanto, en los elementos estructurales de los tejidos musculares hay una gran cantidad de mitocondrias e inclusiones tróficas (gotas de lípidos, gránulos de glucógeno) que contienen sustratos, fuentes de energía. Dado que la contracción muscular ocurre con la participación de iones de calcio, las estructuras que llevan a cabo su acumulación y secreción están bien desarrolladas en las células y fibras musculares, el retículo endoplásmico agranular. (retículo sarcoplásmico), caveola.
Clasificación de tejido muscularbasado en signos de su (a) estructura y función (clasificación morfofuncional)y (b) origen (clasificación histogenética).
Clasificación morfofuncional del tejido muscular. destacados tejido muscular estriado (estriado)y tejido muscular lisoEl tejido muscular estriado está formado por elementos estructurales (células, fibras), que tienen una estriación transversal debido a la disposición mutua ordenada especial de los miofilamentos de actina y miosina en ellos. Los tejidos musculares estriados incluyen esqueléticoy tejido muscular cardíacoEl tejido muscular liso consiste en células que no tienen estriación transversal. El tipo más común de este tejido es el tejido del músculo liso, que forma parte de la pared de varios órganos (bronquios, estómago, intestinos, útero, trompas de Falopio, uréteres, vejiga y vasos sanguíneos).
Clasificación histogenética del tejido muscular. identifica tres tipos principales de tejido muscular: somático(tejido muscular esquelético) celómica(tejido muscular cardíaco) y mesenquimales(tejido muscular liso de los órganos internos), así como dos adicionales: células mioepiteliales(células contráctiles epiteliales modificadas en las secciones terminales y pequeños conductos excretores de algunas glándulas) y elementos minerales(células contráctiles de origen neural en el iris del ojo).
Tejido muscular estriado esquelético (estriado)en su masa excede cualquier otro tejido corporal y es el tejido muscular más común del cuerpo humano. Proporciona movimiento del cuerpo y sus partes en el espacio y mantiene la postura (parte del aparato locomotor), forma músculos oculomotores, músculos de la pared de la cavidad oral, lengua, faringe y laringe. El tejido muscular estriado visceral no esquelético, que se encuentra en el tercio superior del esófago, es parte de los esfínteres externos anales y uretrales.
miocito esquelético muscular
El tejido muscular estriado esquelético se desarrolla en el período embrionario desde miotomossomitas que dan lugar a una división activa mioblastos- celdas que están dispuestas en cadenas y se fusionan entre sí en la región de los extremos con la formación túbulos musculares (miotúbulos)convirtiéndose en fibras muscularesTales estructuras formadas por un solo citoplasma gigante y numerosos núcleos se conocen tradicionalmente en la literatura rusa como simpatistas(en este caso - myosymplasts),sin embargo, este término está ausente en la terminología internacional aceptada. Algunos mioblastos no se fusionan con otros, ubicados en la superficie de las fibras y dando lugar a myosatellitocytes- células pequeñas, que son elementos cambiales del tejido del músculo esquelético. Tejido muscular esquelético formado en haces fibras musculares estriadas, que son sus unidades estructurales y funcionales.
Fibras musculares el tejido del músculo esquelético son formaciones cilíndricas de longitud variable (de milímetros a 10-30 cm). Su diámetro también varía ampliamente dependiendo de la pertenencia a un tipo y músculo en particular, estado funcional, grado de carga funcional, estado nutricional y otros factores. En los músculos, las fibras musculares forman haces en los que se encuentran en paralelo y, al deformarse entre sí, a menudo adquieren una forma multifacética irregular, que se ve especialmente claramente en las secciones transversales. Entre las fibras musculares hay capas delgadas de fibras sueltas. tejido conectivoportando vasos y nervios - endomisioLa estriación transversal de las fibras del músculo esquelético debido a la alternancia de la oscuridad. discos anisotrópicos (bandas A)y brillante discos isotrópicos (bandasYo) Cada disco isotrópico está cortado por la mitad por una delgada y oscura línea Z - telefragm. Los núcleos de la fibra muscular, relativamente livianos, con 1-2 nucleolos, diploides, ovales, aplanados, se encuentran en su periferia debajo del sarcolema y se encuentran a lo largo de la fibra. Fuera del sarcolema está cubierto de espeso membrana basalen el que se tejen las fibras reticulares.
Células de miosatélites (células de miosatélites) - pequeñas células aplanadas ubicadas en impresiones poco profundas del sarcolema de la fibra muscular y cubiertas con una membrana basal común (ver Fig. 88). El núcleo del miosatélite es denso, relativamente grande, los orgánulos son pequeños y pocos. Estas células se activan cuando las fibras musculares se dañan y proporcionan su regeneración reparadora. Al fusionarse con el resto de la fibra con una carga aumentada, las células de los microsatélites están involucradas en su hipertrofia.
Miofibrillas forman el aparato contráctil de la fibra muscular, se ubican en el sarcoplasma a lo largo de su longitud, ocupando la parte central, y se revelan claramente en las secciones transversales de las fibras en forma de pequeños puntos.
Las miofibrillas tienen su propia estriación transversal, y están tan ordenadas en la fibra muscular que los discos isotrópicos y anisotrópicos de diferentes miofibrillas coinciden, causando la estriación transversal de toda la fibra. Cada miofibril está formado por miles de estructuras repetidas interconectadas secuencialmente: sarcómeros.
Sarcomere (myomer)es una unidad estructural y funcional de miofibrillas y representa su porción ubicada entre dos teloperms (líneas Z).Incluye un disco anisotrópico y dos mitades de discos isotrópicos, la mitad de cada lado. Sarcómero formado por un sistema ordenado grueso (miosina)y miofilamentos delgados (actín).Los miofilamentos gruesos están asociados con mesofragmo (línea M)y concentrado en un disco anisotrópico,
y miofilamentos delgados unidos a telogramas (líneas Z),forman discos isotrópicos y penetran parcialmente en el disco anisotrópico entre filamentos gruesos hasta la luz tira Hen el centro del disco anisotrópico.
En el músculo, como en otros tejidos, se distinguen dos tipos de regeneración: fisiológica y reparadora. La regeneración fisiológica se manifiesta en forma de hipertrofia de las fibras musculares, que se expresa en un aumento en su grosor e incluso longitud, un aumento en el número de orgánulos, principalmente miofibrillas, y un aumento en el número de núcleos, que finalmente se manifiesta en un aumento en la capacidad funcional de las fibras musculares. Usando el método de radioisótopos, se descubrió que un aumento en el número de núcleos en las fibras musculares en condiciones de hipertrofia se logra dividiendo las células de los miosatélites y la posterior entrada de las células hijas en el miosymplast.
Se logra un aumento en el número de miofibrillas a través de la síntesis de las proteínas de actina y miosina por los ribosomas libres y el posterior ensamblaje de estas proteínas en miofilamentos de actina y miosina en paralelo con los filamentos de sarcómero correspondientes. Como resultado de esto, al principio hay un engrosamiento de las miofibrillas, y luego su escisión y la formación de miofibrillas hijas. Además, la formación de nuevos miofilamentos de actina y miosina es posible no paralela, sino a tope de las miofibrillas anteriores, logrando así su alargamiento. El retículo sarcoplásmico y los túbulos T en la fibra hipertrófica se forman debido a la proliferación de los elementos anteriores. En ciertos tipos entrenamiento muscular se puede formar un tipo predominantemente rojo de fibra muscular (en styers) o un tipo blanco de fibra muscular (en sprinters). La hipertrofia de las fibras musculares relacionada con la edad se manifiesta intensamente con el inicio de la actividad motora del cuerpo (1-2 años), que se debe principalmente al aumento de la estimulación nerviosa. A la edad senil, así como en condiciones de baja carga muscular, se produce atrofia de orgánulos especiales y generales, adelgazamiento de las fibras musculares y una disminución de su capacidad funcional.
La regeneración reparadora se desarrolla después del daño a las fibras musculares. Además, el método de regeneración depende del tamaño del defecto. Con un daño significativo en toda la fibra muscular, los miosatélites en el área del daño y en áreas adyacentes se desinhiben, proliferan intensamente y luego migran al área del defecto de la fibra muscular, donde se alinean para formar un miotubo. La posterior diferenciación del miotubo conduce a la finalización del defecto y la restauración de la integridad de la fibra muscular. En condiciones de un pequeño defecto en la fibra muscular en sus extremos, debido a la regeneración de los orgánulos intracelulares, se forman riñones musculares que crecen uno hacia el otro y luego se fusionan, lo que lleva al cierre del defecto. Sin embargo, la regeneración reparadora y la restauración de la integridad de las fibras musculares pueden llevarse a cabo bajo ciertas condiciones: en primer lugar, con la inervación motora conservada de las fibras musculares, y en segundo lugar, si los elementos del tejido conectivo (fibroblastos) no caen en el área del daño. De lo contrario, se desarrolla una cicatriz del tejido conectivo en el sitio del defecto de la fibra muscular.
El científico soviético A.N. Student demostró la posibilidad de autotrasplante de tejido del músculo esquelético e incluso de músculos enteros bajo ciertas condiciones:
· Rectificación mecánica del tejido muscular del trasplante, con el objetivo de desinhibir las células satélite y su posterior proliferación;
· La colocación de tejido triturado en el lecho fascial;
· Dobladillo de la fibra nerviosa motora al injerto triturado;
· La presencia de movimientos contráctiles de antagonistas y sinergistas musculares.
Anatómicamente, los recién nacidos tienen todos los músculos esqueléticos, pero con respecto al peso corporal representan solo el 23% (en un adulto 44%). La cantidad de fibra muscular en los músculos es la misma que en un adulto. Sin embargo, la microestructura de las fibras musculares es diferente: las fibras son de menor diámetro, tienen más núcleos. A medida que crece, hay un engrosamiento y alargamiento de las fibras. Esto se debe al engrosamiento de las miofibrillas, empujando el núcleo hacia la periferia. El tamaño de las fibras musculares se estabiliza en 20 años.
Los músculos en los niños son más elásticos que en los adultos. Es decir acortar más rápido al contraer y alargar al relajarse. La excitabilidad y labilidad de los músculos de los recién nacidos es menor que la de los adultos, pero aumenta con la edad. En los recién nacidos, incluso en un sueño, los músculos están en un estado de tono. El desarrollo de varios grupos musculares ocurre de manera desigual. A los 4-5 años, los músculos del antebrazo están más desarrollados, los músculos de la mano se retrasan en el desarrollo. El calentamiento acelerado de los músculos de la mano ocurre en 6-7 años. Además, los extensores se desarrollan más lentamente que los flexores. Con la edad, la proporción del tono muscular cambia. En la primera infancia, aumento del tono de los músculos de la mano, extensores de la cadera, etc. gradualmente la distribución del tono se normaliza.
El corazón como órgano se caracteriza por la capacidad de regenerarse mediante hipertrofia regenerativa, en la que se restaura la masa del órgano, pero la forma permanece alterada. Se observa un fenómeno similar después del infarto de miocardio, cuando la masa del corazón puede recuperarse como un todo, mientras que en el sitio del daño se forma una cicatriz de tejido conectivo, pero el órgano hipertrofia, es decir. El formulario está roto. No solo se produce un aumento en el tamaño de los cardiomiocitos, sino también proliferación principalmente en las aurículas y las orejas del corazón.
Anteriormente se creía que la diferenciación de los cardiomiocitos es un proceso irreversible asociado con la pérdida completa de la capacidad de estas células para dividirse. Pero en el nivel actual, numerosos datos muestran que los cardiomiocitos diferenciados son capaces de síntesis de ADN y mitosis. En los trabajos de investigación de P.P. Rumyantsev y sus estudiantes mostraron que después de un infarto de miocardio ventricular izquierdo experimental, el 60-70% de los cardiomiocitos auriculares regresan al ciclo celular, el número de células poliploides aumenta, pero esto no compensa el daño al miocardio.
Se descubrió que los cardiomiocitos son capaces de división mitótica (incluidas las células del sistema de conducción). En el miocardio del corazón, hay especialmente muchas células poliploides mononucleares con un contenido de ADN de 16-32 veces, pero también hay cardiomiocitos binucleares (13-14%), principalmente octoploides.
En el proceso de regeneración del tejido muscular cardíaco, los cardiomiocitos participan en el proceso de hiperplasia e hipertrofia, su ploidía aumenta, pero el nivel de proliferación de las células del tejido conectivo en el área del daño es 20-40 veces mayor. En los fibroblastos, la síntesis de colágeno se activa, como resultado de lo cual se repara cicatrizando el defecto. La presentación biológica de tal reacción adaptativa del tejido conectivo se explica por la importancia vital del órgano cardíaco, ya que un retraso en el cierre del defecto puede conducir a la muerte.
Se creía que en los recién nacidos, y posiblemente en la primera infancia, cuando aún son capaces de dividir los cardiomiocitos, los procesos regenerativos van acompañados de un aumento en el número de cardiomiocitos. Además, en adultos, la regeneración fisiológica se lleva a cabo en el miocardio principalmente por regeneración intracelular, sin aumentar el número de células, es decir. En el miocardio de un adulto, no hay proliferación de cardiomiocitos. Pero recientemente, se ha obtenido evidencia de que en un corazón humano sano, 14 de un millón de miocitos se encuentran en un estado de mitosis, lo que resulta en una citotomía, es decir. El número de células no es significativo, sino que aumenta.
El uso de métodos modernos de biología celular en estudios clínicos y experimentales nos permitió dilucidar los mecanismos celulares y moleculares del daño y la regeneración del miocardio. De particular interés es la evidencia de que en las regiones perinecróticas y en el corazón funcionalmente congestionado, se lleva a cabo la síntesis de proteínas y péptidos mioarcardiales embrionarios, así como las proteínas sintetizadas durante el ciclo celular. Esto confirma la similitud entre los mecanismos de regeneración y la ontogénesis normal.
También resultó que los cardiomiocitos diferenciados en cultivo son capaces de división mitótica activa, lo que puede explicarse no por una pérdida completa, sino por una supresión de la capacidad de los cardiomiocitos para regresar al ciclo celular.
Una tarea importante de la cardiología teórica y práctica es el desarrollo de formas de estimular la restauración del miocardio dañado, es decir. inducción de la regeneración miocárdica y reducción de la cicatriz del tejido conectivo. Una de las áreas de investigación ofrece la posibilidad de transferir genes reguladores que convierten los fibroblastos de cicatriz en mioblastos o la transfección en cardiomiocitos de genes que controlan el crecimiento de nuevas células. Otra dirección es la transferencia al área de daño de las células esqueléticas y miocárdicas fetales, que podrían estar involucradas en la restauración del músculo cardíaco. También hay experimentos sobre trasplante de músculo esquelético en el corazón, que muestran la formación de áreas de tejido contraído en el miocardio y mejoran los índices funcionales del miocardio. El tratamiento con el uso de factores de crecimiento que tienen efectos tanto directos como indirectos sobre el miocardio dañado, por ejemplo, la mejora de la angiogénesis, puede ser prometedor.
Tejido muscular liso
Tres grupos de tejidos musculares lisos (o no estriados) se distinguen por su origen: mesenquimatoso, epidérmico y neural.
Tejido muscular de origen mesenquimatoso.
Histogénesis Las células madre y las células progenitoras de tejido muscular liso, que ya están determinadas, migran a los lugares de colocación de órganos. Diferenciando, sintetizan los componentes de la matriz y el colágeno de la membrana basal, así como la elastina. En las células definitivas (miocitos), la capacidad sintética se reduce, pero no desaparece por completo.
La unidad estructural y funcional del tejido muscular liso o no estriado es una célula muscular lisa, o un miocito liso es una célula en forma de huso de 20-500 μm de largo, 5-8 μm de ancho. El núcleo de la célula tiene forma de barra, ubicado en su parte central. Cuando el miocito se contrae, su núcleo se dobla e incluso se retuerce. Los orgánulos de importancia general, entre los cuales hay muchas mitocondrias, se concentran en el citoplasma cerca de los polos del núcleo. El aparato de Golgi y el retículo endoplásmico granular están poco desarrollados, lo que indica una baja actividad de las funciones sintéticas. Los ribosomas se encuentran principalmente en libertad.
Los filamentos de actina forman una red tridimensional en el citoplasma, alargada principalmente longitudinalmente, o más bien oblicuamente longitudinalmente. Los extremos de los filamentos están unidos entre sí y al plasmolemma con proteínas de reticulación especiales. Estas áreas son claramente visibles en las micrografías electrónicas como cuerpos densos.
Los filamentos de miosina están en un estado despolimerizado. Los monómeros de miosina se encuentran junto a los filamentos de actina. La señal de contracción generalmente llega a través de las fibras nerviosas. El mediador, que se destaca de sus terminales, cambia el estado del plasmolema. Forma intrusiones: caveola, en la que se concentran iones de calcio. Las caveolas se atan hacia el citoplasma en forma de burbujas (aquí se libera calcio de las burbujas). Esto implica tanto la polimerización de la miosina como la interacción de la miosina con la actina. Los filamentos de actina se desplazan uno hacia el otro, se juntan puntos densos, la fuerza se transmite a la membrana plasmática y se acorta toda la célula. Cuando las señales del sistema nervioso cesan, los iones de calcio son evacuados de las cavidades, la miosina se despolimeriza y las miofibrillas se descomponen. Por lo tanto, los complejos de actina-miosina existen en los miocitos lisos solo durante el período de contracción.
Los miocitos lisos se encuentran sin espacios intercelulares notables y están separados por una membrana basal. En ciertas áreas, se forman "ventanas" en él, por lo que el plasmolema de los miocitos vecinos converge. Aquí se forman nexos, y no solo surgen enlaces mecánicos, sino también metabólicos entre las células. En la parte superior de las "cubiertas" de la membrana basal entre los miocitos hay fibras elásticas y reticulares, que combinan las células en un solo complejo tisular. Las fibras reticulares penetran en las grietas en los extremos de los miocitos, se fijan allí y transmiten la fuerza de la contracción celular a toda su asociación.
Regeneracion La regeneración fisiológica del tejido muscular liso se manifiesta en condiciones de mayores cargas funcionales. Esto se ve más claramente en la membrana muscular del útero durante el embarazo. Dicha regeneración se lleva a cabo no tanto a nivel de tejido como a nivel celular: los miocitos crecen, los procesos sintéticos se activan en el citoplasma, aumenta el número de miofilamentos (hipertrofia celular activa). Sin embargo, la proliferación celular (es decir, hiperplasia) no está excluida.
Como parte de los órganos, los miocitos se combinan en haces, entre los cuales hay capas delgadas de tejido conectivo. Las fibras reticulares y elásticas que rodean a los miocitos se tejen en estas capas. Los vasos sanguíneos y las fibras nerviosas pasan a través de las capas intermedias. Los terminales de este último no terminan directamente en los miocitos, sino entre ellos. Por lo tanto, después de la llegada de un impulso nervioso, el mediador se propaga difusamente, excitando muchas células a la vez. El tejido muscular liso de origen mesenquimatoso está representado principalmente en las paredes de los vasos sanguíneos y en muchos órganos internos tubulares, y también forma músculos pequeños individuales.
El tejido muscular liso en órganos específicos tiene propiedades funcionales desiguales. Esto se debe al hecho de que en la superficie de los órganos hay diferentes receptores para sustancias biológicamente activas específicas. Por lo tanto, para muchas drogas, su reacción no es la misma.
Tejido muscular liso de origen epidérmico.
Las células mioepiteliales se desarrollan a partir del germen epidérmico. Se encuentran en las glándulas sudoríparas, mamarias, salivales y lagrimales y tienen precursores comunes con células secretoras glandulares. Las células mioepiteliales se unen directamente al epitelio propiamente dicho y tienen una membrana basal común con ellas. Durante la regeneración, esas y otras células se restauran a partir de precursores comunes poco diferenciados. La mayoría de las células mioepiteliales tienen forma de estrella. Estas células a menudo se llaman en forma de canasta: sus procesos abarcan las secciones finales y los pequeños conductos de las glándulas. El núcleo y los orgánulos de importancia general se encuentran en el cuerpo de la célula, y el aparato contráctil, organizado, como en las células del tejido muscular del tipo mesenquimatoso, se ubica en los procesos.
Tejido muscular liso de origen neural.
Los miocitos de este tejido se desarrollan a partir de células del rudimento neural como parte de la pared interna de la copa óptica. Los cuerpos de estas células se encuentran en el epitelio de la superficie posterior del iris. Cada uno de ellos tiene un proceso que entra en el grosor del iris y se encuentra paralelo a su superficie. En el proceso hay un aparato contráctil, organizado de la misma manera que en todos los miocitos lisos. Dependiendo de la dirección de los procesos (perpendicular o paralelo al borde de la pupila), los miocitos forman dos músculos: una pupila que se estrecha y se expande.
Conclusión
Como ya se señaló, el tejido muscular es un grupo de tejidos corporales de diversos orígenes, unidos por el principio de contractilidad: estriado (esquelético y cardíaco), liso, así como tejidos contráctiles especializados: epitelial-muscular y neuroglial, que forma parte del iris del ojo.
El tejido del músculo esquelético estriado surge de los miotomas que forman los elementos del mesodermo segmentado: somitas.
El tejido muscular liso de humanos y vertebrados se desarrolla como parte de los derivados del mesénquima, así como del tejido interno. Sin embargo, todos los tejidos musculares se caracterizan por un aislamiento similar en el embrión embrionario en forma de células en forma de huso: células formadoras de músculo o mioblastos.
La contracción de la fibra muscular implica acortar miofibrillas dentro de cada sarcómero. Los filamentos gruesos (miosina) y delgados (actina), en un estado relajado, conectados solo por las secciones finales, en el momento de la contracción realizan movimientos deslizantes uno hacia el otro. La liberación de energía necesaria para reducir la energía se produce como resultado de la conversión de ATP a ADP bajo la influencia de la miosina. La actividad enzimática de la miosina se manifiesta bajo la condición de contenido óptimo de Ca2 +, que se acumula en el retículo sarcoplásmico.
Referencias
1. Histología. Editado por Yu.I. Afanasyeva, N.A. Yurina M .: "Medicina", 1999
2. R. Eckert, D. Rendel, J. Augustine "Fisiología Animal" - 1 vol. M .: "Mir", 1981
3. K.P. Ryabov "Histología con los fundamentos de la embriología" Minsk: "Escuela superior", 1990
4. Histología. Editado por Ulumbekov, prof. Yu.A. Chelysheva M .: 1998
5. Histología. Editado por V.G. Eliseeva M .: "Medicina", 1983
Publicado en Allbest.ru
...Documentos similares
Características estructurales del tejido muscular. El estudio del mecanismo de contracción muscular y el aparato de transmisión de la excitación. Histogénesis y regeneración del tejido muscular. Los principios de los cardiomiocitos contráctiles, conductores y secretores del tejido muscular cardíaco.
hoja de trucos añadida el 14/11/2010
El estudio de los tipos y funciones de diversos tejidos humanos. Tareas de la ciencia de la histología, que estudia la estructura de los tejidos de los organismos vivos. Características estructurales del tejido epitelial, nervioso, muscular y tejidos del ambiente interno (conectivo, esquelético y fluido).
presentación agregada el 08/11/2013
El estudio de las características estructurales de los tejidos animales, el funcionamiento y las variedades. Realización de un estudio de un rasgo característico de la estructura del tejido conectivo y nervioso. La estructura del epitelio plano, cúbico, ciliado y glandular. Tipos de tejido muscular.
presentación agregada el 08/02/2015
Características generales y características relacionadas con la edad del tejido cartilaginoso. Tipos de cartílago y tejido óseo. Características generales y características del tejido óseo relacionadas con la edad. Características de la estructura del tejido muscular en la infancia y la vejez. Tejido muscular esquelético.
presentación, agregada 07/02/2016
Clasificación de tejidos, tipos de tejidos epiteliales, su estructura y funciones. Función de soporte, trófica y protectora de los tejidos conectivos. Las funciones de los tejidos nerviosos y musculares. El concepto de órganos y sistemas de órganos, sus diferencias individuales, de género y edad.
resumen, agregado 09/11/2009
Tejido epitelial, su capacidad regenerativa. Tejidos conectivos involucrados en el mantenimiento de la homeostasis del ambiente interno. Células de refugio y linfa. Tejido muscular estriado y cardíaco. Las funciones de las células nerviosas y los tejidos de los organismos animales.
resumen, agregado el 16 de enero de 2015
El estudio de los tipos de tejido animal, así como las funciones que realizan. Características estructurales de los grupos de tejido epitelial, conectivo, muscular y nervioso. Determinar la ubicación de cada grupo y la importancia para la vida del animal.
presentación, agregada 18/10/2013
Concepto general y tipos de vibraciones. Caracterización de los procesos de tensión (compresión), cizallamiento, flexión, torsión. Las propiedades mecánicas del hueso y el tejido vascular. La especificidad del tejido muscular, los principales modos de trabajo muscular: isométrico e isotónico.
trabajo de control, agregado 19/03/2014
La célula como la principal unidad estructural del cuerpo. Descripción de su estructura, propiedades vitales y químicas. La estructura y funciones del tejido epitelial y conectivo, muscular y nervioso. Órganos y una lista de sistemas de órganos humanos, su propósito y funciones.
presentación agregada el 19/04/2012
Fisiología y bioquímica de la actividad muscular como un componente importante del metabolismo en el cuerpo. Tipos de tejido muscular y, en consecuencia, músculos que difieren en la estructura de las fibras musculares, la naturaleza de la inervación. El efecto de la actividad física de intensidad variable.
Capítulo 9. TEJIDOS MUSCULARES
Capítulo 9. TEJIDOS MUSCULARES
Tejido muscular (textus muscularis)llamados telas que son diferentes en estructura y origen, pero similares en capacidad de reducción. Proporcionan movimiento en el espacio del cuerpo como un todo, sus partes y el movimiento de los órganos dentro del cuerpo (corazón, lengua, intestinos, etc.).
Las células de muchos tejidos poseen la propiedad de contraerse con un cambio de forma, pero en los tejidos musculares esta capacidad se convierte en la función principal.
9.1. CARACTERÍSTICAS MORFOFUNCIONALES GENERALES Y CLASIFICACIÓN
Los principales signos morfológicos de los elementos del tejido muscular son la forma alargada, la presencia de miofibrillas y miofilamentos ubicados longitudinalmente, orgánulos especiales que proporcionan contractilidad, la ubicación de las mitocondrias cerca de elementos contráctiles, la presencia de inclusiones de glucógeno, lípidos y mioglobina.
Los orgánulos contráctiles especiales, miofilamentos o miofibrillas, proporcionan la contracción que ocurre cuando dos proteínas fibrilares principales, la actina y la miosina, interactúan con ellas, con la participación obligatoria de iones de calcio. Las mitocondrias proporcionan energía a estos procesos. El stock de fuentes de energía está formado por glucógeno y lípidos. La mioglobina es una proteína que proporciona la unión del oxígeno y la creación de su reserva en el momento de la contracción muscular, cuando se comprimen los vasos sanguíneos (el flujo de oxígeno disminuye bruscamente).
ClasificaciónLa clasificación del tejido muscular se basa en dos principios: morfofuncional e histogenético. De acuerdo con el principio morfológico y funcional, dependiendo de la estructura de los orgánulos de contracción, el tejido muscular se divide en dos subgrupos.
Primer subgrupo- tejido muscular estriado (estriado) (Textus muscularis striatus).En el citoplasma de sus elementos, miosina filamen-
estás constantemente polimerizado, forma miofibrillas constantemente existentes con filamentos de actina. Estos últimos están organizados en complejos característicos: sarcómerosEn las miofibrillas vecinas, las subunidades estructurales de los sarcómeros se encuentran al mismo nivel y crean estriación cruzada
Segundo subgrupo- tejido muscular liso (no estriado) (Textus muscularis no estriado).Estos tejidos se caracterizan por el hecho de que, sin contracción, los filamentos de miosina se despolimerizan. En presencia de iones de calcio, polimerizan e interactúan con los filamentos de actina. Las miofibrillas formadas en este caso no tienen una estriación transversal: con colores especiales están representadas por filamentos (lisos) coloreados uniformemente a lo largo de toda la longitud.
De acuerdo con el principio histogenético, dependiendo de las fuentes de desarrollo (primordios embrionarios), los tejidos musculares y los elementos musculares se dividen en: somático (miotomo), celómico (de la placa mioepicárdica de la hoja de esplenototomo visceral), mesenquimatoso (del germen desmal en el mesénquima (neural), neural tubo neural), epidérmico (del ectodermo cutáneo y de la placa precordal).
9.2. TEJIDOS MUSCULARES RAYADOS
Hay dos variedades principales de tejidos estriados (estriados): esquelético (miotomo) y cardíaco (celómico).
9.2.1. Tejido muscular esquelético
HistogénesisLa fuente del desarrollo de los elementos del tejido muscular estriado esquelético (somático) (textus muscularis striatus sceletalis)son células madre de miotomas: promioblastos. Algunos de ellos se diferencian en el acto y participan en la formación de los llamados músculos autóctonos. Otras células migran de miotomas al mesénquima. Ya están determinados, aunque externamente no difieren de otras células mesenquimatosas. Su diferenciación continúa en los sitios donde se colocan otros músculos del cuerpo. Durante la diferenciación, surgen dos líneas celulares. Las células de una de las líneas se fusionan, formando simpáticos alargados: tubos musculares (miotubos). En ellos, la diferenciación de orgánulos especiales - miofibrillas (Fig. 9.1). En este momento, se observa un retículo endoplásmico granular bien desarrollado en los miotubos. Las miofibrillas se ubican primero debajo del plasmolema y luego llenan la mayoría de los miotubos. Los núcleos, por el contrario, de los departamentos centrales se trasladan a la periferia. Los centros celulares y los microtúbulos desaparecen por completo. Endo granular
Fig. 9.1.Histogénesis del tejido del músculo esquelético (según A. A. Klishov):
pero- promioblastos; b- miosymplast; en- tubo muscular; g- músculo maduro
fibra 1 - miosatélite; 2 - núcleo miosymplast; 3 - miofibrillas
la red de plasma se reduce significativamente. Tales estructuras definitivas se denominan miosimplastias.
Las células de la otra línea permanecen independientes y se diferencian en células miosatélite. Estas células se encuentran en la superficie de los miosimmplastos. Las células de miosatélites, que se multiplican, se fusionan con miosimmplastos, participando así en la creación de un sistema nuclear óptimo
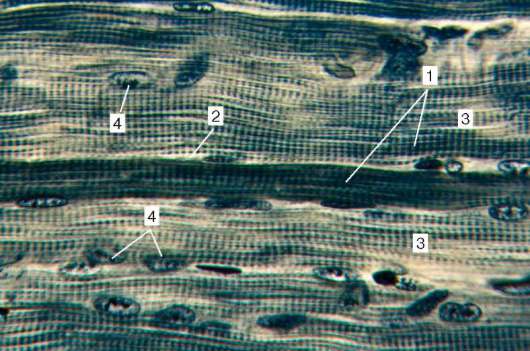
Fig. 9.2.La estructura del tejido muscular estriado (micrografía):
1 - fibras musculares; 2 - sarcolema; 3 - sarcoplasma y miofibrillas; 4 - núcleos
miosimplastia Colorear - Hematoxilina De Hierro
relación sarcoplásmica necesaria para la síntesis de proteínas ejemplares específicas.
La estructuraLa unidad estructural principal del tejido del músculo esquelético es la fibra muscular, que consiste en miosimplastias y miostelocitos, cubierta con una membrana basal común (fig. 9.2-9.4). La longitud de toda la fibra se puede medir en centímetros con un grosor de 50-100 micras. El complejo, que consiste en el plasmolema del miosimplastio y la membrana basal, se llama sarcolema.
La estructura del miosymplast.Myosymplast tiene muchos núcleos oblongos ubicados directamente debajo del plasmolema. Su número en un simplast puede alcanzar varias decenas de miles (ver Fig. 9.2). Los orgánulos de importancia general, el complejo de Golgi y pequeños fragmentos del retículo endoplásmico agranular, se encuentran en los polos de los núcleos. Las miofibrillas llenan la parte principal del miosimmplast y se ubican longitudinalmente (ver Fig. 9.3).
Sarcomere- unidad estructural de miofibrillas. Cada miofibril tiene discos transversales oscuros y claros con refracción desigual (discos A anisotrópicos e discos isotrópicos I). Cada miofibril está rodeado por asas de un retículo endoplásmico agranular, un retículo sarcoplásmico, ubicado longitudinalmente y anastomosándose entre sí. Los sarcómeros vecinos tienen una estructura de borde común: la línea Z (Fig. 9.5). Está construido en forma de una red de moléculas de proteínas fibrilares, entre las cuales la alfa-actinina juega un papel importante. Los extremos de los filamentos de actina están conectados a esta red. Desde las líneas Z vecinas, los filamentos de actina se dirigen al centro del sarcómero, pero no alcanzan su centro. Filamentos de actina combinados con la línea Z y filamentos de miosina
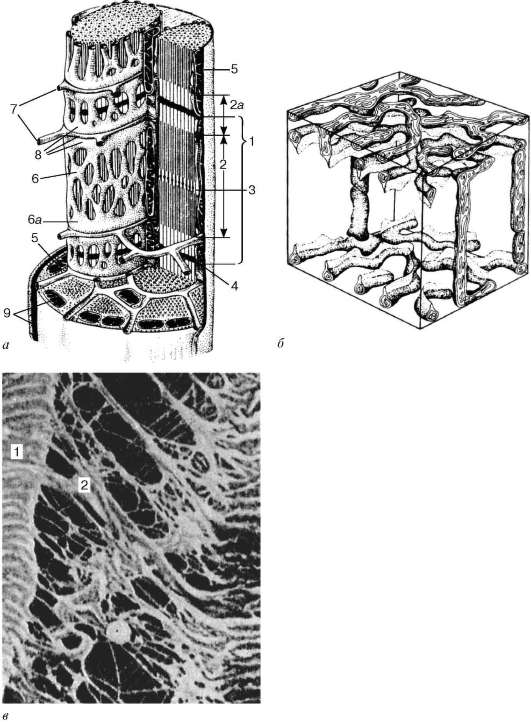
Fig. 9.3.Esquema de la estructura ultramicroscópica del miosymplast (según R.V. Krstic, con cambios) (a): 1 - sarcómero; 2 - disco anisotrópico (carril A); 2a - disco isotrópico (carril I); 3 - línea M (mesofragma) en el medio del disco anisotrópico; 4 - línea Z (telofragma) en el medio de un disco isotrópico; 5 - mitocondrias; 6 - retículo sarcoplásmico; 6a - tanque final; 7 - túbulo transversal (tubo en T); 8 - tríada; 9 - sarcolema; b- La disposición espacial de las mitocondrias en el symplast. Los planos superior e inferior de la figura están limitados por la sarco-medida del disco anisotrópico (según L. E. Bakeeva, V. P. Skulachev, Yu. S. Chentsov); en- endomisio Micrografía electrónica de barrido, aumento 2600 (preparación de Yu. A. Khoroshkov): 1 - fibras musculares; 2 - fibrillas de colágeno
moléculas de nebulina fibrilares inextensibles. En el medio del disco oscuro del sarcómero hay una red hecha de miomiosina. Forma una línea M en sección transversal. En los nodos de esta línea M, los extremos de los filamentos de miosina están fijos. Sus otros extremos están dirigidos hacia las líneas Z y
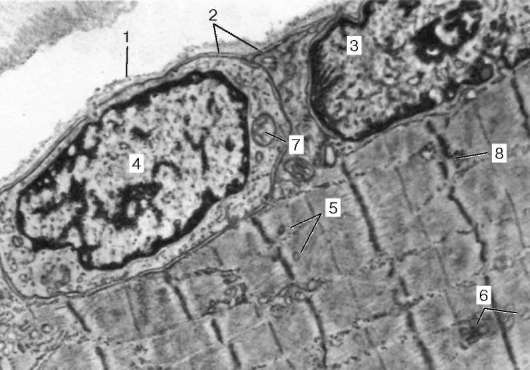
Fig. 9.4.El área de superficie del miosimplastio y el miosatélite. Micrografía electrónica, aumento de 10.000 (preparación por V. L. Goryachkina y S. L. Kuznetsova): 1 - membrana basal; 2 - plasmolemma; 3 - el núcleo del miosymplast; 4 - el núcleo del mios-telitocito; 5 - miofibrillas; 6 - túbulos de la red endoplásmica agranular (sarcoplasmática); 7 - mitocondrias; 8 - glucógeno
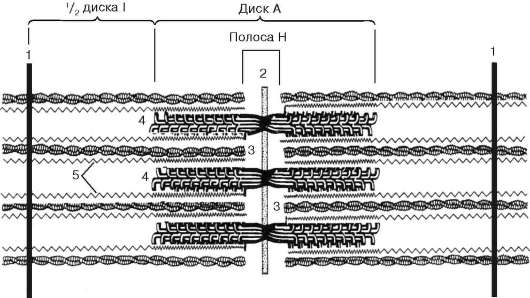
Fig. 9.5.Sarcómero (diagrama):
1 - línea Z; 2 - línea M; 3 - filamentos de actina; 4 - filamentos de miosina; 5 - moléculas de titina fibrilar (según B. Alberts, D. Bray, J. Lewis et al., Según enmendada)
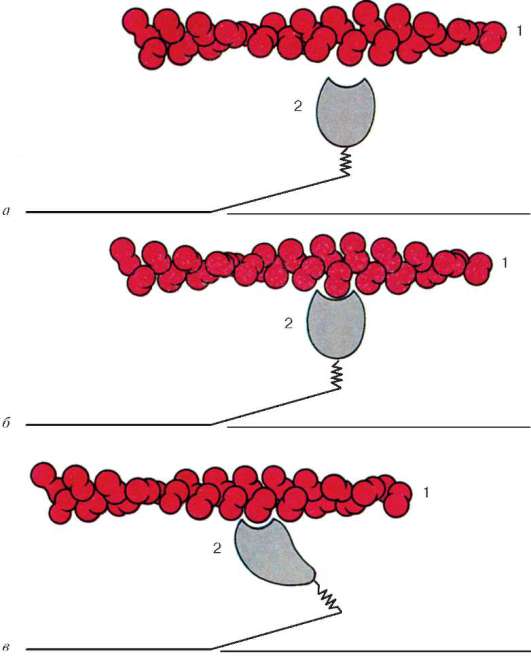
Fig. 9.6.Cambios conformacionales que implican un desplazamiento mutuo de filamentos de actina y miosina:
a-b- cambios sucesivos en las relaciones espaciales. 1 - actina; 2 - la cabeza de la molécula de miosina (según B. Alberts, D. Bray, J. Lewis et al., Según enmendada)
ubicado entre los filamentos de actina, pero tampoco alcanza las líneas Z en sí. Al mismo tiempo, estos extremos se fijan con respecto a las líneas Z mediante moléculas de titina de proteína gigante extensibles.
Las moléculas de miosina tienen una cola larga y dos cabezas en uno de sus extremos. Con un aumento en la concentración de iones de calcio en el área de unión de las cabezas (porción de bisagra), la molécula cambia su configuración (Fig. 9.6). Al mismo tiempo (dado que la actina se encuentra entre los filamentos de miosina), las cabezas de miosina se unen a la actina (con la participación de proteínas auxiliares: tropomiosina y troponina). Luego, la cabeza de miosina se inclina y tira de la molécula de actina junto con ella hacia la línea M. Las líneas Z convergen, el sarcómero se acorta.
Las redes de alfa-actinina de las líneas Z de las miofibrillas vecinas están interconectadas por filamentos intermedios. Se acercan a la superficie interna del plasmolemma y se fijan en su capa cortical, de modo que los sarcómeros de todas las miofibrillas se encuentran en el mismo nivel. Esto, cuando se observa bajo un microscopio, crea la impresión de la estriación transversal de toda la fibra.
La fuente de iones de calcio son los tanques del retículo endoplásmico agranular. Se alargan a lo largo de las miofibrillas cerca de cada sarcómero y forman un retículo sarcoplásmico. Es en él que los iones de calcio se acumulan cuando el miosymplast está en un estado relajado. Al nivel de las líneas Z (en anfibios) o en el borde de los discos A e I (en mamíferos), los túbulos de la red cambian de dirección y son transversales, formando tanques terminales o laterales (L) extendidos.
Desde la superficie hasta la profundidad del miosymplast, el plasmolema forma túbulos largos que se extienden transversalmente (túbulos en T) al nivel de los límites entre los discos oscuros y claros. Cuando el miosymplast recibe una señal sobre el comienzo de la contracción, se mueve a lo largo del plasmolema en forma de potencial de acción y se extiende a la membrana del tubo en T. Dado que esta membrana está cerca de las membranas del retículo sarcoplásmico, el estado de este último cambia, el calcio se libera de los tanques de la red e interactúa con los complejos de actina-miosina (se contraen). Cuando el potencial de acción desaparece, el calcio se acumula nuevamente en los túbulos de la red y se detiene la contracción de las miofibrillas. Se necesita energía para desarrollar un esfuerzo de reducción. Se libera convirtiendo ATP a ADP. El papel de ATPasa lo realiza la miosina. La fuente de ATP es principalmente las mitocondrias, por lo que se encuentran directamente entre las miofibrillas.
Las inclusiones de mioglobina y glucógeno desempeñan un papel importante en la actividad de los miosimmplastos. El glucógeno sirve como fuente de energía, necesaria no solo para realizar trabajo muscular, sino también para mantener el equilibrio térmico de todo el organismo. La mioglobina se une al oxígeno cuando el músculo está relajado y la sangre fluye libremente a través de los pequeños vasos sanguíneos. Durante la contracción muscular, los vasos sanguíneos se comprimen y el oxígeno almacenado se libera y participa en reacciones bioquímicas.
Células miosatélite.Estas células mal diferenciadas son una fuente de regeneración del tejido muscular. Son adyacentes a la superficie del miosimmplast, de modo que sus plasmolemas están en contacto (ver Fig. 9.1, 9.4). Las células de los miosatélites son mononucleares, sus núcleos oscuros son ovales y más pequeños que en los simpáticos. Poseen todos los orgánulos de importancia general (incluido el centro celular).
Tipos de fibras musculares.Los diferentes músculos (como los órganos) funcionan en diferentes condiciones biomecánicas. Por lo tanto, las fibras musculares en la composición de diferentes músculos tienen diferente fuerza, velocidad y duración de la contracción, así como también fatiga. La actividad de las enzimas en ellos es diferente, y se presentan en diversas formas isoméricas. El contenido de enzimas respiratorias, glicolíticas y oxidativas, también es diferente en ellas.
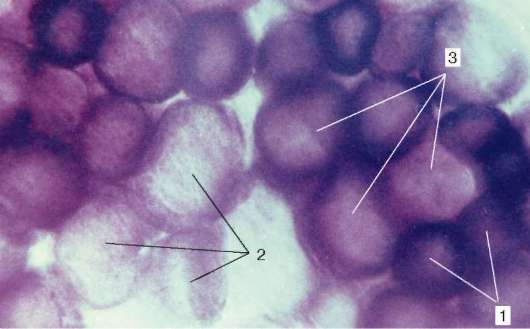
Fig. 9.7La actividad de la succinato deshidrogenasa en fibras musculares de diversos tipos (fármaco V. F. Chetvergova, procesamiento según Nakhlas et al.): 1 - alto; 2 - bajo; 3 - mediano
Por la proporción de miofibrillas, mitocondrias y mioglobina, se distinguen las fibras blancas, rojas e intermedias. Por características funcionales, las fibras musculares se dividen en rápidas, lentas e intermedias, lo que está determinado por la organización molecular de la miosina. Entre sus isoformas, hay dos principales: "rápido" y "lento". Al organizar las reacciones histoquímicas, se identifican por la actividad de ATPasa. La actividad enzimática respiratoria también se correlaciona con estas propiedades. Por lo general, los procesos glucolíticos predominan en las fibras rápidas, son ricos en glucógeno, tienen menos mioglobina y, por lo tanto, se llaman blancos. Por el contrario, en las fibras lentas, la actividad de las enzimas oxidantes es mayor; son más ricas en mioglobina y se ven más rojas.
Junto con el blanco y el rojo, hay fibras intermedias. En la mayoría de los músculos esqueléticos, las fibras de diferentes tipos histoquímicos son de mosaico (Fig. 9.7).
Las propiedades de las fibras musculares cambian con las cargas cambiantes: deportivas, profesionales, así como en condiciones extremas (ingravidez). Cuando vuelve a las actividades normales, dichos cambios son reversibles. En algunas enfermedades (atrofia muscular, distrofia, consecuencias de la denervación), las fibras musculares con diferentes propiedades iniciales cambian de manera diferente. Esto le permite aclarar el diagnóstico, para lo cual se examinan muestras de biopsia de músculos esqueléticos.
RegeneracionLos núcleos de miosimmplastos no pueden dividirse, ya que no hay centros celulares en el sarcoplasma. Los elementos cambiales sirven células de miosatélites.Mientras el cuerpo crece, se dividen, y las células hijas se fusionan con miosimmplastos. Al final del crecimiento, la reproducción de las células de los miosatélites se desvanece. Después del daño a la fibra muscular en cierta medida desde el sitio de la lesión, se destruye y su fragmento
usted está fagocitado por los macrófagos. La restauración del tejido se lleva a cabo mediante dos mecanismos: hipertrofia compensatoria del síntoma mismo y proliferación de células de miosatélites. En el ejemplo, se activan el retículo endoplásmico granular y el complejo de Golgi. Se produce la síntesis de las sustancias necesarias para la restauración del sarcoplasma y las miofibrillas, así como el ensamblaje de las membranas, de modo que se restablezca la integridad de la lemma plasmática. En este caso, el extremo dañado del miosymplast se engrosa, formando un riñón muscular. Las células de los miosatélites que sobreviven cerca de la lesión se dividen. Algunos de ellos migran al riñón muscular y se integran en él, mientras que otros se fusionan (al igual que los mioblastos durante la histogénesis) y forman nuevos miotubos que se convierten en fibras musculares.
9.2.2. Músculo esquelético como órgano
La transmisión de los esfuerzos de contracción al esqueleto se realiza a través de los tendones o uniendo los músculos directamente al periostio. Al final de cada fibra muscular, el plasmolemma forma protuberancias profundas y estrechas. Las fibras de colágeno delgadas penetran desde el tendón o el periostio. Estos últimos están trenzados en espiral con fibras reticulares. Los extremos de las fibras se dirigen a la membrana basal, entran en ella, giran hacia atrás y, al salir, vuelven a clasificar las fibras de colágeno del tejido conectivo.
Entre las fibras musculares hay capas delgadas de tejido conectivo fibroso suelto. endomisioLas fibras de colágeno de la hoja externa de la membrana basal se tejen en ella (ver Fig. 9.3, c), lo que ayuda a combinar los esfuerzos al tiempo que reduce los miosimmplastos. Las capas más gruesas de tejido conectivo laxo rodean varias fibras musculares, formando perimisioy dividiendo el músculo en paquetes. Varios paquetes se combinan en grupos más grandes, separados por capas de tejido conectivo más gruesas. El tejido conectivo que rodea la superficie del músculo se llama epimisio
VascularizaciónLas arterias ingresan al músculo y se extienden a través de las capas de tejido conectivo, disminuyendo gradualmente. Las ramas de quinto y sexto orden forman arteriolas en el perimisio. Los capilares se encuentran en el endomisio. Van a lo largo de las fibras musculares, anastomosándose entre sí. Las vénulas, las venas y los vasos linfáticos pasan cerca de los vasos de entrega. Como de costumbre, hay muchos mastocitos cerca de los vasos involucrados en la regulación de la permeabilidad de la pared vascular.
InervaciónEl músculo reveló fibras nerviosas eferentes (motoras), aferentes (sensibles), así como fibras nerviosas vegetativas no mielinizadas. Scion célula nerviosatrayendo impulso de la neurona motora médula espinalramas en perimisia. Cada una de sus ramas penetra a través de la membrana basal y forma terminales en la superficie del simplast en el plasmolemma, participando en la organización de la llamada placa motora (véase el capítulo 10, Fig. 10.18). Al ingreso
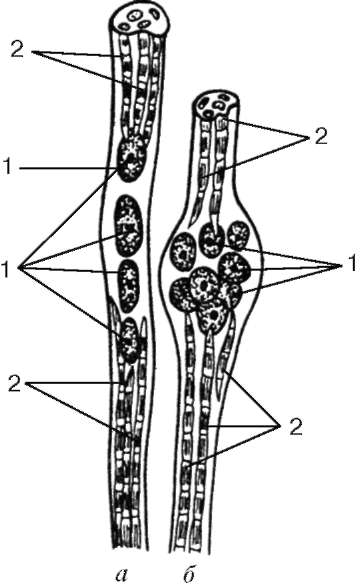
Fig. 9.8Un fragmento de un huso muscular que contiene fibras musculares con una cadena nuclear (a) y con una bolsa nuclear (b) (esquema según G. S. Katinas): 1 - núcleos; 2 - miofibrillas (no se muestran orgánulos de importancia general)
la acetilcolina se libera desde los terminales del impulso nervioso, un mediador que causa excitación (potencial de acción), que se propaga desde aquí a través del plasmolema miosimmplast.
Entonces, cada fibra muscular está inervada de forma independiente y está rodeada por una red de hemocapilares, formando un complejo llamado myon
El grupo de fibras musculares inervadas por una neurona motora se llama unidad neuromuscular.Las fibras musculares que pertenecen a una unidad neuromuscular no se encuentran cerca, sino que se encuentran en mosaico entre las fibras que pertenecen a otras unidades.
Las terminaciones nerviosas sensibles no se encuentran en las fibras musculares de trabajo (extrafusales), sino que están conectadas con fibras musculares especializadas en los llamados músculos musculares.
tenakh (con fibras musculares intrafusales), que se encuentran en el perimisio.
Fibras musculares intrafusales.Las fibras musculares intrafusales de los husos son mucho más delgadas que los trabajadores. Hay dos tipos: fibra con una bolsa nuclear y fibra con una cadena nuclear (Fig. 9.8). Los núcleos en esos y en otros están redondeados y ubicados en el grosor del symplast, y no en su superficie. En fibras con una bolsa nuclear, el núcleo de la forma más simple se agrupa en su parte media engrosada. En las fibras con una cadena nuclear en la parte media del sistema simple, no se forma un engrosamiento, los núcleos se encuentran aquí uno tras otro. Los orgánulos de importancia general se encuentran cerca de grupos de núcleos.
Las miofibrillas se encuentran en los extremos de los simpáticos. La fibra de sarcolema se conecta a la cápsula del huso neuromuscular, que consiste en un tejido conectivo fibroso denso. Cada fibra muscular del huso está entrelazada en espiral con un terminal de la fibra nerviosa sensorial. Como resultado de la reducción o relajación de las fibras musculares activas, la tensión de la cápsula del tejido conectivo del huso cambia, respectivamente, cambia el tono de las fibras musculares intrafusales. Como resultado de esto, las terminaciones nerviosas sensibles se excitan a su alrededor y aparecen impulsos nerviosos aferentes en el área terminal. Cada miosimmplast también tiene su propia placa motora. Es por eso que las fibras musculares intrafusales están constantemente en tensión, adaptándose a la longitud del abdomen muscular en su conjunto.
9.2.3. Tejido muscular del corazón
Histogénesis y tipos de células.Fuentes de desarrollo del tejido muscular estriado cardíaco (textus muscularis striatus cardiacus)- secciones simétricas del splanchnotome de la hoja visceral en la parte cervical del embrión - placas mioepicárdicas.De estos, las células del mesotelioma epicardio también se diferencian. Las células originales del tejido del músculo cardíaco: explosiones de cardiomyo- Se caracteriza por una serie de signos: las células están aplanadas, contienen un núcleo grande, un citoplasma ligero, pobre en ribosomas y mitocondrias. En el futuro, se desarrolla el complejo de Golgi, una red granular de endo-plasma. Las estructuras fibrilares se encuentran en los cardiomioblastos, pero no hay miofibrillas. Las células tienen un alto potencial proliferativo.
Después de una serie de ciclos mitóticos, los cardiomioblastos se diferencian en cardiomiocitosen el que comienza la sarcomerogénesis (fig. 9.9). En el citoplasma de los cardiomiocitos, aumenta el número de polisomas y túbulos del retículo endoplásmico granular, se acumulan los gránulos de glucógeno y aumenta el volumen del complejo de actomiosina. Los cardiomiocitos se contraen, pero no pierden la capacidad de proliferar y diferenciarse aún más. El desarrollo del aparato contráctil en los últimos períodos embrionarios y postnatales ocurre al agregar nuevos sarcómeros y estratificar miofilamentos recién sintetizados.
La diferenciación de los cardiomiocitos se acompaña de un aumento en el número de mitocondrias, su distribución en los polos de los núcleos y entre las miofibrillas y continúa en paralelo con la especialización de las superficies de contacto de las células. Los cardiomiocitos forman contactos de "extremo a extremo", de "extremo a lado" para formar fibras musculares cardíacas y, en general, el tejido es una estructura tipo red. Algunos cardiomiocitos en las primeras etapas de la cardiomiogénesis son secretoras contráctiles. Posteriormente, como resultado de la diferenciación divergente, surgen miocitos "oscuros" (contráctiles) y "claros" (conductores), en los que los gránulos secretores desaparecen, mientras permanecen en los miocitos auriculares. Entonces se forma el diferencial de cardiomiocitos endocrinos. Estas células contienen un núcleo ubicado en el centro con cromatina dispersa, uno o dos nucleolos. En el citoplasma, un retículo endoplásmico granular, dictiosomas del complejo de Golgi, está bien desarrollado, en estrecha conexión con los elementos de los cuales hay numerosos gránulos secretores con un diámetro de aproximadamente 2 μm, que contiene material denso en electrones. Posteriormente, los gránulos secretores se encuentran debajo del sarcolema y se excretan en el espacio intercelular por exocitosis.
En general, durante la histogénesis surgen cinco tipos de cardiomiocitos: de trabajo (contráctil), sinusal (marcapasos), de transición, conductivo y secretorio. Cardiomiocitos de trabajo (contráctiles)forman sus cadenas (Fig. 9.10). Son ellos, cuando se acortan, los que proporcionan la fuerza de contracción de todo el músculo cardíaco. Los cardiomiocitos de trabajo son capaces de
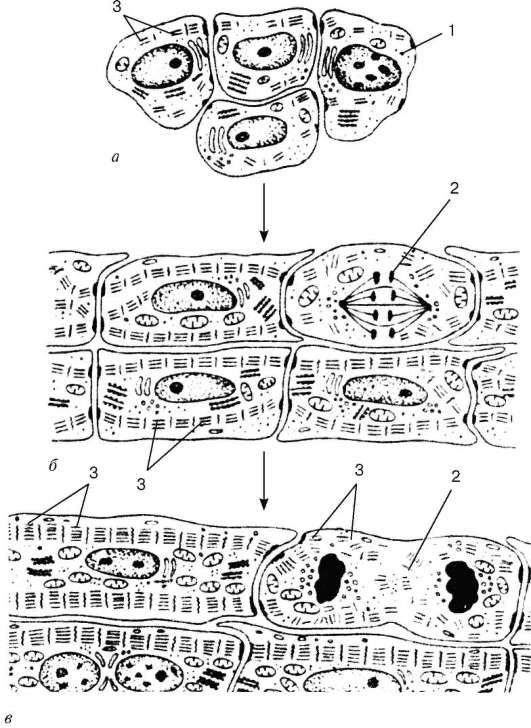
Fig. 9.9Histogénesis del tejido muscular cardíaco (esquema según P.P. Rumyantsev): pero- cardiomiocitos en la pared del tubo cardíaco; b -cardiomiocitos en embriogénesis tardía; en- cardiomiocitos en el período postanatal. 1 - cardiomiocito; 2 - cardiomiocito que se divide mitóticamente; 3 - miofilamentos y miofibrillas
transmitir señales de control entre sí. Cardiomiocitos sinusales (marcapasos)capaz de cambiar automáticamente en cierto ritmo el estado de contracción a un estado de relajación. Las células perciben señales de control de las fibras nerviosas, en respuesta a las cuales cambian el ritmo de la actividad contráctil. Los cardiomiocitos sinusales (marcapasos) transmiten señales de control cardiomiocitos transitorios,y el último a los cardiomiocitos conductores y de trabajo. Conducir cardiomiocitosforman cadenas de células conectadas por sus extremos y se ubican debajo del endo
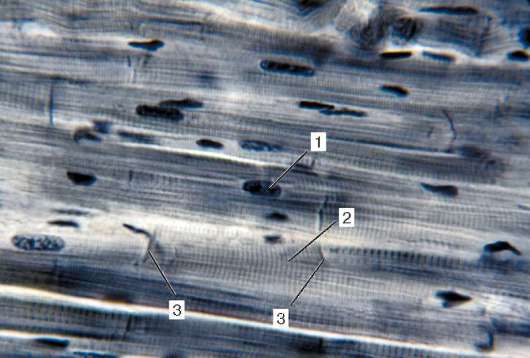
Fig. 9.10.La estructura del tejido muscular cardíaco (micrografía). Coloración - hematoxilina de hierro:
1 - núcleo de cardiomiocitos; 2 - una cadena de cardiomiocitos; 3 - insertar ruedas
cardom. La primera célula de la cadena recibe señales de control de los cardiomiocitos sinusales y las pasa a otros cardiomiocitos conductores. Las células que cierran la cadena transmiten una señal a través de los cardiomiocitos de transición a los trabajadores. Cardiomiocitos Secretoresrealizar una función especial Producen la hormona peptídica cardiodilatina, que circula en la sangre como cardionatrina, provoca una reducción de los miocitos lisos en las arteriolas, un aumento en el flujo sanguíneo renal y acelera la filtración glomerular y la excreción de sodio. Todos los cardiomiocitos están cubiertos con una membrana basal.
La estructura de los cardiomiocitos contráctiles (de trabajo).Las células tienen una forma alargada (100-150 μm), casi cilíndrica. Sus extremos están conectados entre sí, de modo que las cadenas celulares forman las llamadas fibras funcionales (hasta 20 micras de espesor). En el área de los contactos celulares, se forman los llamados discos de inserción (Fig. 9.10). Los cardiomiocitos pueden ramificarse y formar una red espacial. Sus superficies están cubiertas con una membrana basal, en la que se tejen fibras reticulares y de colágeno desde el exterior. El núcleo de un cardiomiocito (a veces dos de ellos) es ovalado y se encuentra en la parte central de la célula. En los polos del núcleo, se concentran pocos orgánulos de importancia general, con la excepción del retículo endoplásmico agranular y las mitocondrias.
Los orgánulos especiales que proporcionan contracción se llaman miofibrillas. Están débilmente aislados unos de otros, se pueden dividir. Su estructura es similar a la estructura de las miofibrillas del miosymplast de la fibra del músculo esquelético. Cada mitocondria se encuentra en todo el sarcómero. Los tubos en T ubicados al nivel de la línea Z se dirigen desde la superficie de la membrana plasmática hacia la profundidad del cardiomiocito. Sus membranas se unen
en contacto con las membranas de la red endoplásmica lisa (sarcoplasmática). Los bucles de estos últimos están alargados a lo largo de la superficie de las miofibrillas y tienen extensiones laterales (sistemas L), que forman, junto con los tubos en T, una tríada o diada (Fig. 9.11, a). En el citoplasma hay inclusiones de glucógeno y lípidos, especialmente muchas inclusiones de mioglobina. El mecanismo de contracción de los cardiomiocitos es el mismo que en miosymplast.
Organización de los cardiomiocitos en el tejido.Los cardiomiocitos están conectados entre sí por el tipo de "extremo a extremo". Aquí se forman discos de inserción: estas áreas parecen placas delgadas con un aumento promedio del microscopio óptico. De hecho, los extremos de los cardiomiocitos tienen una superficie irregular, por lo que las protuberancias de una célula entran en los canales de otra. Las secciones transversales de las protuberancias de las células adyacentes están interconectadas por interdigitaciones y desmosomas (Fig. 9.11, b).
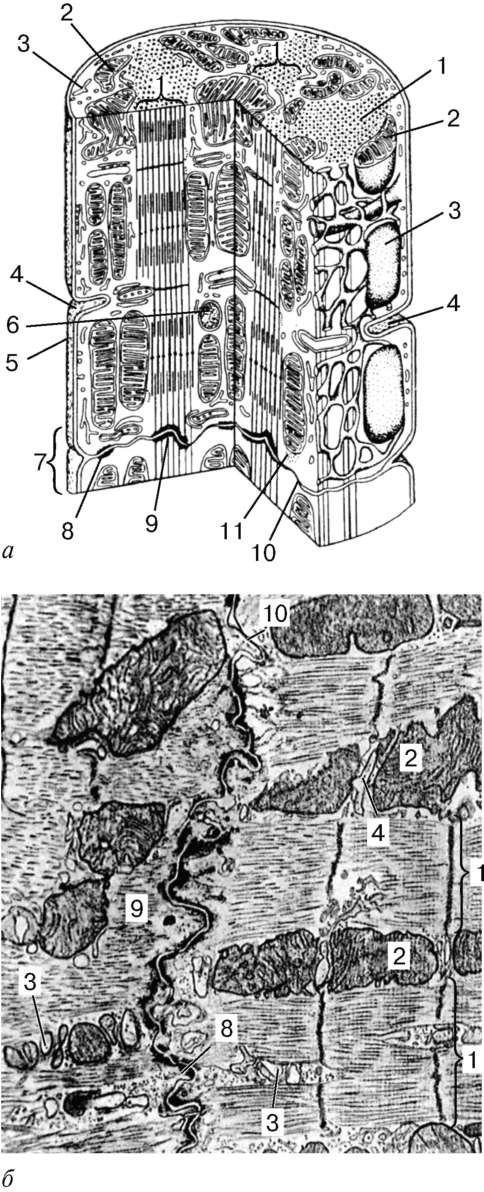
Fig. 9.11.La estructura del cardiomiocito: pero- esquema (según Yu. I. Afanasyev y V. L. Goryachkina); b- Micrografía electrónica del disco de inserción. Un aumento de 20,000.1 - miofibrillas; 2 - mitocondrias; 3 - red sarcotubular; 4 - tubos en T; 5 - membrana basal; 6 - lisosoma; 7 - insertar disco; 8 - desmosoma; 9 - zona de unión de miofibrillas; 10 - contactos ranurados; 11 - glucógeno
A cada desmosoma desde el lado del citoplasma, se acerca miofibril la, que se fija al final en el complejo de la placa desmo. Por lo tanto, mientras se reduce el deseo de un cardiomiocito se transmite a otro. Las superficies laterales de las protuberancias de los cardiomiocitos están unidas por nexos (uniones huecas). Esto crea enlaces metabólicos entre ellos y asegura la sincronización de las contracciones.
RegeneracionEn la histogénesis del tejido muscular cardíaco, el cambium no ocurre. Por lo tanto, la regeneración de tejidos se realiza sobre la base de procesos hiperplásicos intracelulares. Al mismo tiempo, el proceso de poliploidía es característico de los cardiomiocitos de mamíferos, primates y humanos.
nación Por ejemplo, en los monos, hasta el 50% de los cardiomiocitos diferenciados terminalmente en el núcleo se convierten en tetra y octoploide. Los cardiomiocitos poliploides se producen debido a la mitosis acitocinética, que conduce a la multinucleación. Bajo las condiciones de la patología del sistema cardiovascular humano (reumatismo, defectos cardíacos congénitos, infarto de miocardio, etc.), la regeneración intracelular, la poliploidización de los núcleos y la aparición de cardiomiocitos multinucleados juegan un papel importante en la compensación del daño a los cardiomiocitos.
9.3. TELAS MUSCULARES LISAS
Hay tres grupos de tejido muscular liso (no estriado) (textus muscularis no estriado)y células: células mesenquimales, neurales y mioepiteliales.
9.3.1. Tejido muscular de origen mesenquimatoso.
HistogénesisEste tejido se divide en dos tipos: visceral y vascular. En la histogénesis embrionaria, incluso la microscopía electrónica es difícil de distinguir entre los progenitores de fibroblastos mesenquimales y los miocitos lisos. En los miocitos lisos poco diferenciados, se desarrolla un retículo endoplásmico granular, complejo de Golgi. Los filamentos delgados están orientados a lo largo del eje largo de la célula. A medida que se desarrollan, aumentan el tamaño de las células y la cantidad de filamentos en el citoplasma. Gradualmente, el volumen del citoplasma ocupado por filamentos contráctiles aumenta, su ubicación en el citoplasma se vuelve cada vez más ordenada. La actividad proliferativa de los miocitos lisos en la miogénesis se reduce gradualmente. Esto ocurre como resultado de un aumento en la duración del ciclo celular, la salida de las células del ciclo reproductivo y la transición a un estado diferenciado. Al diferenciarse, sintetizan los componentes de la matriz intercelular, el colágeno de la membrana basal y la elastina. En las células definitivas (miocitos), la capacidad sintética se reduce, pero no desaparece por completo.
La estructura y el funcionamiento de las células.Un miocito liso es una célula en forma de huso de 20-500 micras de largo, 5-8 micras de ancho. El núcleo tiene forma de varilla, ubicado en su parte central. Cuando el miocito se contrae, su núcleo se dobla e incluso se retuerce (fig. 9.12–9.14).
La estructura de los miocitos lisos definitivos (leiomiocitos) que forman los órganos internos y las paredes de los vasos sanguíneos tiene mucho en común, pero al mismo tiempo se caracteriza por el heteromorfismo. Entonces, en las paredes de las venas y arterias se encuentran ovoides, en forma de huso, procesan miocitos con una longitud de 10-40 micras, que a veces alcanzan hasta 140 micras. El mio suave más largo
los citos alcanzan hasta 500 micras en la pared uterina. El diámetro de los miocitos varía de 2 a 20 micras. Dependiendo de la naturaleza de los procesos biosintéticos intracelulares, se distinguen los miocitos contráctiles y secretores. Los primeros están especializados en la función de contracción, pero al mismo tiempo retienen la actividad secretora.
Los miocitos secretores en su ultraestructura se parecen a los fibroblastos, pero contienen en su citoplasma haces de miofilamentos delgados ubicados en la periferia de la célula. El complejo de Golgi, el retículo endoplásmico granular, muchas mitocondrias, gránulos de glucógeno, ribosomas libres y polisomas están bien desarrollados en el citoplasma. Por el grado de madurez, tales células se clasifican como poco diferenciadas. Los filamentos de actina en el citoplasma forman una red tridimensional, alargada principalmente longitudinalmente. Los extremos de los filamentos están unidos entre sí y al plasmolemma con proteínas de reticulación especiales. Estas áreas son claramente visibles en las micrografías electrónicas como cuerpos densos. Los monómeros de miosina se encuentran junto a los filamentos de actina. El plasmolema forma intrusiones: caveolas, en las que se concentran los iones de calcio. La señal de contracción generalmente llega a través de las fibras nerviosas. El mediador, que se destaca de sus términos, cambia la permeabilidad del plasmolemma. Se liberan iones de calcio, lo que implica tanto la polimerización de la miosina como la interacción de la miosina con la actina.
Hay una retracción de actina-miofilamentos entre miosi
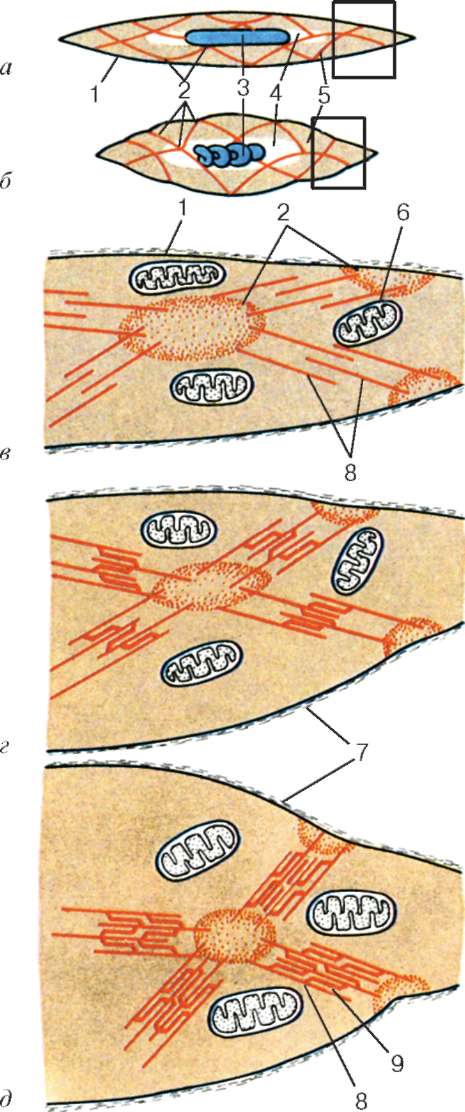
Fig. 9.12.La estructura de un miocito liso (esquema):
un en- cuando se relaja; b d- a la mayor reducción; g- con reducción incompleta; wd- imágenes ampliadas de secciones enmarcadas en fragmentos peroy b. 1 - plasmolemma; 2 - cuerpos densos; 3 - núcleo; 4 - endoplasma; 5 - complejos contráctiles; 6 - mitocondrias; 7 - membrana basal; 8 - miofilamentos de actina (delgados); 9 - miofilamentos de miosina (gruesos)
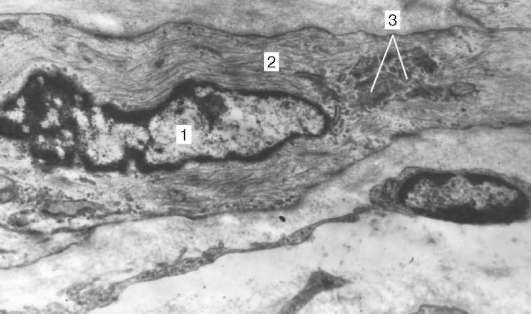
Fig. 9.13.Ultraestructura de miocitos lisos diferenciadores en la pared del bronquio:
1 - núcleo; 2 - citoplasma con miofilamentos; 3 - Complejo de Golgi, un aumento de 35 000 (fármaco A. L. Zashikhina)
se juntan puntos nuevos y densos, la fuerza se transmite al plasmolema y se acorta toda la célula (ver Fig. 9.12). Cuando cesan las señales del sistema nervioso, los iones de calcio se mueven del citoplasma a la caveola y los túbulos del retículo endoplásmico, la miosina se despolimeriza y las miofibrillas se rompen. La contracción se detiene. Por lo tanto, los complejos de actinomiosina existen en los miocitos lisos solo durante el período de contracción en presencia de iones de calcio libres en el citoplasma.
Los miocitos están rodeados por una membrana basal. En ciertas áreas, se forman "ventanas" en él, por lo que el plasmolema de los miocitos vecinos converge. Aquí se forman nexos, y no solo surgen enlaces mecánicos, sino también metabólicos entre las células. En la parte superior de las "cubiertas" de la membrana basal entre los miocitos hay fibras elásticas y reticulares, que combinan las células en un solo complejo tisular. Los miocitos lisos sintetizan proteoglucanos, glicoproteínas, procolágeno, pro-elastina, a partir de los cuales se forman colágeno y fibras elásticas y el componente amorfo de la matriz intercelular. La interacción de los miocitos se lleva a cabo mediante puentes citoplasmáticos, protuberancias mutuas, nexos, desmosomas, secciones de contactos de membrana de las superficies de los miocitos.
RegeneracionEl tejido muscular liso de las especies viscerales y vasculares tiene una sensibilidad significativa a los efectos de factores extremos. En los miocitos activados, el nivel de procesos biosintéticos aumenta, cuya expresión morfológica es la síntesis de proteínas contráctiles, agrandamiento e hipercromatosis del núcleo, hipertrofia del nucleolo, un aumento en la relación citoplasmática nuclear, un aumento en el número de ribosomas y polisomas libres, y

Fig. 9.14.La estructura del tejido muscular liso (esquema volumétrico) (según R.V. Krstic, con cambios):
1 - miocitos lisos fusiformes; 2 - citoplasma de miocitos; 3 - núcleos de miocitos; 4 - plasmolemma; 5 - membrana basal; 6 - vesículas pinocitóticas superficiales; 7 - conexiones intercelulares; 8 - terminación nerviosa; 9 - fibrillas de colágeno; 10 - microfilamentos
ción de enzimas, fosforilación aeróbica y anaeróbica, transporte de membrana. La regeneración celular se lleva a cabo tanto por las células diferenciadas con la capacidad de ingresar al ciclo mitótico, como por la activación de elementos cambiales (miocitos de pequeño volumen). Bajo la acción de varios factores dañinos, se observa una transformación fenotípica de los miocitos contráctiles en secretores. Esta transformación a menudo se observa con daño a la íntima vascular, la formación de hiperplasia de la íntima con el desarrollo de aterosclerosis.
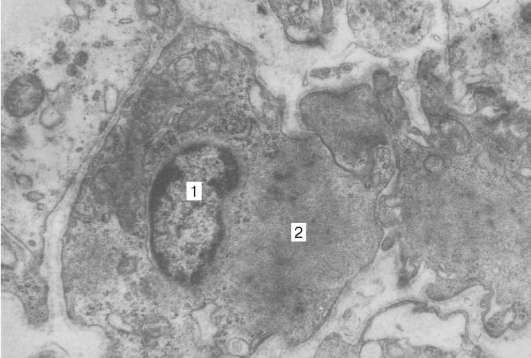
Fig. 9.15.Ultraestructura del miopigmentocito (fármaco N. N. Sarbaeva): 1 - núcleo; 2 - miofilamentos, un aumento de 6000
9.3.2. Tejido muscular mesenquimatoso en órganos
Los miocitos se combinan en paquetes, entre los cuales hay capas delgadas de tejido conectivo. Las fibras reticulares y elásticas que rodean a los miocitos se tejen en estas capas. Los vasos sanguíneos y las fibras nerviosas pasan a través de las capas intermedias. Los terminales de este último no terminan directamente en los miocitos, sino entre ellos. Por lo tanto, después de la llegada de un impulso nervioso, el mediador se propaga difusamente, excitando muchas células a la vez. El tejido muscular liso de origen mesenquimatoso está representado principalmente en las paredes de los vasos sanguíneos y en muchos órganos internos huecos.
El tejido muscular liso en órganos específicos tiene propiedades funcionales desiguales. Esto se debe al hecho de que en la superficie de los órganos hay diferentes receptores para sustancias biológicamente activas específicas. Por lo tanto, para muchas drogas, su reacción no es la misma. Es posible que diferentes propiedades funcionales de los tejidos estén asociadas con una organización molecular específica de los filamentos de actina.
9.3.3. Tejido muscular de origen neural.
El tejido muscular del iris y el cuerpo ciliar es el cuarto tipo de tejido contráctil. Los miocitos de este tejido se desarrollan a partir de células del rudimento neural como parte de la pared interna de la copa óptica. En una fila
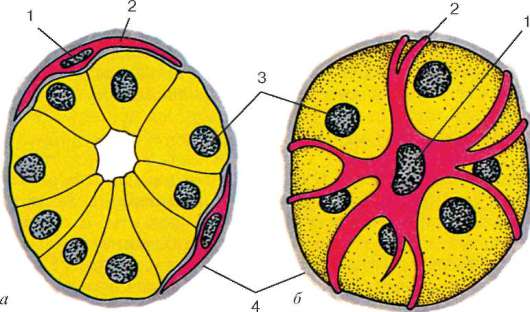
Fig. 9.16.Células mioepiteliales en la sección final de la glándula salival (esquema según G. S. Katinas):
pero- sección transversal; b- Vista desde la superficie. 1 - núcleos de células mioepiteliales; 2 - procesos de células mioepiteliales; 3 - núcleos de células epiteliales secretoras; 4 - membrana basal
los elementos del músculo vertebral del iris exhiben una diferenciación divergente diversa. Por lo tanto, el tejido mioneural en reptiles y aves está representado por fibras multinucleadas estriadas, que son muy similares al músculo esquelético. En mamíferos y humanos, la principal unidad estructural y funcional de los músculos del iris es un miocito mononuclear liso o miopigmentocito. Estos últimos tienen un cuerpo pigmentado que contiene un núcleo, tomado fuera de la parte contráctil en forma de huso (Fig. 9.15).
El citoplasma de las células contiene una gran cantidad de mitocondrias y gránulos de pigmento, que son similares en tamaño y forma a los gránulos del epitelio pigmentario. Los miofilamentos en los miopigmentocitos se dividen en delgados (7 nm) y gruesos (1,5 nm), en tamaño y ubicación se parecen a los miofilamentos de miocitos lisos. Cada miopigmentocito está rodeado por una membrana basal. Cerca de los procesos citoplasmáticos de los miocitos, se encuentran fibras nerviosas libres de mielina. Dependiendo de la dirección de los procesos (perpendicular o paralelo al borde de la pupila), los miocitos forman dos músculos: una pupila que se estrecha y se expande.
RegeneracionAlgunos estudios han demostrado una baja actividad regenerativa después del daño o su ausencia.
9.3.4. Células musculares epidérmicas
Las células mioepiteliales se desarrollan a partir del germen epidérmico. Se encuentran en las glándulas sudoríparas, mamarias, salivales y lagrimales y tienen precursores comunes con sus células secretoras. Mioepitelial
las células están directamente adyacentes al epitelio propiamente dicho y tienen una membrana basal común con ellas. Durante la regeneración, esas y otras células también se restauran a partir de precursores comunes poco diferenciados. La mayoría de las células mioepiteliales tienen forma de estrella. Estas células a menudo se denominan en forma de canasta: sus procesos abarcan las secciones finales y los pequeños conductos de las glándulas (Fig. 9.16). El núcleo y los orgánulos de importancia general se encuentran en el cuerpo de la célula, y el aparato contráctil, organizado como en las células del tejido muscular del tipo mesenquimatoso, se ubica en los procesos.
Preguntas de seguridad
1. Clasificación genética del tejido muscular. Unidades estructurales y funcionales de diferentes tipos de tejido muscular.
2. Tejido estriado del músculo esquelético: desarrollo, estructura, base morfológica de la contracción muscular. Regeneracion
3. Tejido muscular cardíaco estriado: desarrollo, estructura específica de varios tipos de cardiomiocitos, regeneración.
4. Variedades de miocitos lisos: fuentes de desarrollo, topografía en el cuerpo, regeneración.
Histología, embriología, citología: libro de texto / Yu. I. Afanasyev, N. A. Yurina, E. F. Kotovsky y otros. - 6ª ed., Rev. y agregar. - 2012 .-- 800 s. : enfermo
Los músculos del cuerpo humano están formados principalmente por tejido muscular, que consiste en células musculares. Distinguir entre tejido muscular liso y estriado. Tejido muscular liso acumula musculatura, que forma parte de algunos órganos internos, y estriadoforma músculo esquelético Una propiedad común del tejido muscular es su excitabilidad, conductividad y contractilidad.
El tejido muscular estriado se distingue del liso, mayor excitabilidad, conductividad y contractilidad. Las células del músculo estriado tienen un diámetro muy pequeño y una longitud grande (hasta 10-12 cm). En este sentido, se llaman fibras.
Al igual que otras células, las células musculares tienen un protoplasma llamado sarcoplasma(del griego. Sarcos - carne). Membrana de células musculares llamada sarcolema. Dentro de la fibra muscular hay numerosos núcleos y otros componentes de las células.
La composición de las fibras musculares incluye una gran cantidad de fibras aún más delgadas. miofibrillas, que, a su vez, consisten en los hilos más delgados: protofibrillas. Protofibrillas Este es el aparato contráctil de la célula muscular, son proteínas contráctiles especiales. miosina y actina. El mecanismo de las contracciones musculares es un proceso complejo de transformaciones físicas y químicas en la fibra muscular con la participación obligatoria del aparato contráctil. Este mecanismo es activado por un impulso nervioso, y la energía para el proceso de reducción es suministrada por el ácido trifosfórico de adenosina (ATP). En este sentido, una característica de la estructura de las fibras musculares es también una gran cantidad de mitocondrias que proporcionan a la fibra muscular la energía necesaria. La relajación de la fibra muscular, según muchos científicos, se lleva a cabo pasivamente, debido a la elasticidad del sarcolema y el tejido conectivo intramuscular.
La estructura, forma y clasificación del músculo esquelético.
La unidad anatómica de la parte más activa del sistema muscular humano (músculos esqueléticos o estriados) es el músculo esquelético. Músculo esquelético - Este es un órgano formado por tejido muscular estriado, que también contiene tejido conectivo, nervios y vasos sanguíneos.
Cada músculo está rodeado por una especie de "caso" de tejido conectivo: fascia.
En estructura externa Los músculos distinguen entre la cabeza del tendón correspondiente al comienzo del músculo, el abdomen del músculo o el cuerpo formado por fibras musculares, y el extremo del tendón del músculo, o cola, con el que el músculo se une a otro hueso. Típicamente, la cola del músculo es un punto de unión móvil, y el comienzo es fijo. En el proceso de movimiento, sus funciones pueden cambiar: los puntos de movimiento se vuelven inmóviles y viceversa.
La forma de los músculos es muy diversa y depende en gran medida del propósito funcional del músculo. Distinga entre músculos largos, cortos, anchos, circulares y otros. Largo los músculos están ubicados en las extremidades corta - allí, donde el movimiento es pequeño (por ejemplo, entre las vértebras). Los músculos anchos se encuentran principalmente en el cuerpo (músculos del abdomen, la espalda, el pecho). Los músculos circulares están ubicados alrededor de las aberturas del cuerpo y los contraen cuando se contraen. Estos músculos se llaman esfínteres. Si un músculo tiene una cabeza, se llama simple, si dos o más son complejos (por ejemplo, bíceps, tríceps y cuádriceps). Una característica morfológica importante es la ubicación de las fibras musculares. Hay disposición paralela, oblicua, transversal y circular de fibras (en esfínteres). si, con una disposición oblicua de fibras musculares, se unen solo en un lado a los tendones, entonces los músculos se llaman pinnatesi en ambos lados bifurcado.
Funcionalmente, los músculos se pueden dividir en flexores y extensores, rotadores afuera y rotadores en el interior, músculos aductores y descarga. Los músculos también son secretados. sinergistas y músculos antagonistas. Los primeros forman un grupo de músculos, amigos que realizan cualquier movimiento, la contracción de los segundos provoca movimientos opuestos.
Grupos musculares principales
Los músculos esqueléticos de un adulto representan aproximadamente el 40% de su peso corporal. En los recién nacidos y los niños pequeños, los músculos no representan más del 20-25% del peso corporal, y en la vejez hay una disminución gradual de la masa muscular hasta el 25-30% del peso corporal. En total, el cuerpo humano tiene más de 600 músculos.
Por ubicación, es decir en terrenos topográficos, se distinguen los músculos de la espalda, pecho, abdomen, cabeza, cuello, extremidades superiores e inferiores.
Los músculos de la espalda se dividen en superficial y profundo. A músculos superficiales las espaldas incluyen: músculo trapecio, dorsal ancho, músculos romboides (mayor y menor), músculos dentados (superior e inferior).
Estos músculos están involucrados en los movimientos de las extremidades superiores y los cambios de volumen. el cofre (acto de respirar).
Músculos profundos Las espaldas están representadas por numerosos músculos ubicados a lo largo de la columna vertebral. Están unidos principalmente a los procesos de las vértebras y están involucrados en los movimientos de la columna hacia atrás y hacia los lados.
Los músculos del cofre se dividen en músculos que pertenecen a la cintura escapular y miembro superior (músculos pectorales mayor y menor, músculos subclavios y dentados anteriores) , y músculos propios del tórax (músculos intercostales internos externos). Ponen en movimiento y fortalecen las extremidades superiores del cuerpo y participan en la función de la respiración. El diafragma también pertenece a los músculos respiratorios (separa la cavidad torácica de la cavidad abdominal).
Los músculos abdominales están representados por los músculos oblicuo, transverso y recto abdominal externo e interno, así como el músculo cuadrado de la espalda baja.
Los músculos rectos abdominales participan en la flexión hacia adelante del cuerpo, y los músculos oblicuos proporcionan la inclinación de la columna hacia los lados y sus giros junto con el pecho hacia la derecha y hacia la izquierda. Los músculos abdominales no solo participan en los movimientos del tronco y el pecho. Forman una pared cavidad abdominal y, debido a su tono, mantienen sus órganos internos en su posición normal. Al reducirlos, aumentan la presión intraabdominal, formando el llamado abdominal. Los músculos abdominales facilitan las deposiciones (evacuaciones), la micción y la expulsión del feto en las mujeres durante el parto. La contracción de los músculos abdominales también contribuye al movimiento de la sangre en el sistema venoso y a la implementación de movimientos respiratorios, pero también a la formación de una hernia. Con una hernia, se produce una protuberancia de los órganos internos (intestinos, estómago, etc.) desde la cavidad abdominal debajo de la piel.
Entre los músculos de la cabeza, se distinguen los músculos para masticar y los faciales. A masticar músculos aplica temporal, masticatorio, pterigoideo. La contracción de estos músculos provoca movimientos complejos de masticación. mandíbula inferior. Los músculos miméticos difieren de otros músculos del cuerpo humano en que están unidos a la piel de la cara con uno, y a veces dos, de sus extremos. La contracción de estos músculos es el desplazamiento de la piel, la formación de pliegues y arrugas. Esto define expresiones faciales, es decir una u otra expresión facial La manifestación de sensaciones complejas (emociones): alegría, vergüenza, desprecio, pena y dolor, etc. - Está determinado por numerosas combinaciones de contracciones musculares faciales, subordinadas a los impulsos que les llegan desde la corteza cerebral a lo largo del nervio facial. Los músculos circulares del ojo y la boca también son músculos imitadores. Los músculos del cuello inclinan la cabeza, la inclinan y giran. Los músculos unidos al hueso hioides, durante la contracción, cambian la posición de la lengua y la laringe al tragar y emitir sonidos.
Los músculos de la extremidad superior se dividen en músculos. cintura escapular y músculos de la extremidad superior libre. Los músculos de la cintura escapular rodean la articulación del hombro, proporcionando numerosos movimientos en ella. Entre ellos, el más importante es el músculo deltoides. Al contraerse, este músculo dobla el brazo articulación del hombro y lleva su mano a la posición horizontal.
Los músculos de la extremidad superior libre incluyen los músculos del hombro, el antebrazo y la mano.
En el área del hombro al frente hay un grupo muscular: flexoresatrás - extensores. Entre los músculos del grupo anterior, el más importante es el músculo bíceps del hombro (bíceps), y la espalda, el músculo tríceps del hombro.
Los músculos del antebrazo también están representados en la superficie frontal por los flexores, y en la parte posterior por los extensores. Los músculos de la mano se encuentran solo en la superficie palmar. Entre ellos se destacan: un músculo palmar largo y flexores de los dedos. Debido a la presencia de su propio sistema muscular, los dedos, especialmente pulgar, adquieren una mayor movilidad y son capaces de realizar varios movimientos, incluida la máxima flexión y extensión, lo cual es extremadamente importante cuando se trabaja.
Los músculos ubicados en la región del cinturón de la extremidad inferior mueven la pierna hacia adentro articulación de la cadera, así como con una extremidad fija, doblan la columna vertebral en la región lumbar. El grupo muscular anterior incluye un músculo principal: el iliopsoas. Entre el grupo externo posterior de músculos de la cintura pélvica: músculos glúteos grandes, medianos y pequeños.
Miembros inferiores tienen un esqueleto más masivo que los superiores; sus músculos tienen una gran fuerza, pero al mismo tiempo menos variedad y rango de movimiento limitado.
En el muslo al frente se encuentra el músculo sastre más largo del cuerpo humano (hasta 50 cm). Ella dobla la pierna en las articulaciones de la cadera y la rodilla.
El músculo cuádriceps femoral se encuentra más profundo que el músculo sastre, rodeando el fémur desde casi todos los lados.
La función principal de este músculo es la extensión de la articulación de la rodilla. Al estar de pie, el cuádriceps no cede articulación de la rodilla agacharse
En el pie, se distinguen los músculos de la espalda y plantares. Estos músculos están involucrados en la flexión y extensión de los dedos de los pies, así como en su convergencia y reproducción.